

В настоящей главе рассматриваются современные воззрения на физико-химическую организацию и физико-химические свойства клетки. Однако специфика живой клетки такова, что она не исчерпывается только описанием происходящих в ней химических и физических процессов и структурных перестроек. Основой этой специфики и ее характерной чертой является тесная взаимосвязь и взаимообусловленность физических, физико-химических и биохимических процессов с процессами физиологическими, отражающими свойства и реакции клетки как целого. Поэтому современный этап в развитии физиологии клетки характеризуется тесным взаимодействием изучения физико-химических процессов в живой материи в «молекулярном масштабе» и тех процессов, которые являются клеточными реакциями, т. е. реакциями, складывающимися из многообразного переплетения функций и свойств биологических структур клетки.
Физико-химическая организация протоплазмы
Основой клеточной организации, выражением ее сущности, как живого, является протоплазма — основное содержимое любой живой клетки. Протоплазма характеризуется гомогенностью, нерастворимостью в воде, эластичностью, сократимостью, способностью к обратимым изменениям своего состава и вязкости. Эти свойства протоплазмы сохраняются лишь до тех пор, пока это вещество находится в живой интактной клетке.
С физико-химической точки зрения протоплазму можно рассматривать как сложную коллоидную систему, обладающую всеми свойствами и признаками макромолекул в растворе.
Определение коллоидного состояния вещества можно дать, основываясь на следующих признаках: 1) коллоиды не проходят через поры естественных мембран; 2) коллоидные частицы отделены от непрерывной среды активными поверхностями; 3) коллоидные растворы обладают сильным светорассеиванием (эффект Тиндэля); 4) коллоиды отличаются очень малой скоростью диффузии.
Эти общие свойства коллоидов относятся к весьма обширной группе коллоидных систем; в сложной гетерогенной, среде (какой, в частности, является протоплазма) может присутствовать одновременно несколько коллоидных систем.
Важной коллоидной системой являются эмульсии. Характерным свойством эмульсий является то, что дисперсная фаза и дисперсионная среда в них — жидкие, однако они не смешиваются между собой. Примером могут служить капельки жира и масла (микроскопического и субмикроскопического размеров), находящиеся в водной фазе протоплазмы.
Другой важной колоидной системой являются эмульсоиды. Их отличительная черта — взаимное соединение между дисперсионной средой и дисперсной фазой. Стабильность таких коллоидных систем объясняется двумя свойствами: наличием электрического заряда и наличием сил сцепления с дисперсионной средой (образование так называемого сольватационного слоя).
В сольватационном слое молекулы растворителя (например, воды) ориентируются и поляризуются под влиянием электрического заряда. К эмульсоидам относятся растворы белков и некоторых углеводов в воде.
Еще более сложной системой коллоидов являются коацерваты (рис. 1). Коацерваты представляют собой такую коллоидную систему, в которой произошло отмешивание (разделение) однородного коллоидного раствора на два слоя — обедненный и обогащенный коллоидными частицами (Евреинова, 1954). В результате действия поливалентных ионов или взаимодействия двух гидрофильных коллоидов противоположных зарядов возникают так называемые сложные коацерваты (рис. 2). Поддержанию стабильности сложных коацерватов способствует то, что при их образовании происходит увеличение сил притяжения между положительно и отрицательно заряженными частицами коллоида.
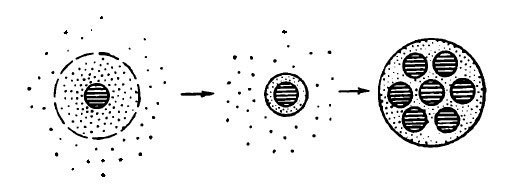
Рис. 1. Образование коацерватов
(Bungenberg бе Jong, 1956)
По современным воззрениям, протоплазма построена по типу сложных коацерватов. Белки протоплазмы представляют собой (большей частью конъюгированные белки, т. е. соединения простых белков с другими «веществами (нуклеиновыми кислотами, углеводами, высшими жирными кислотами и т. д.). При соединении с белком эти вещества образуют сложные коацерваты.
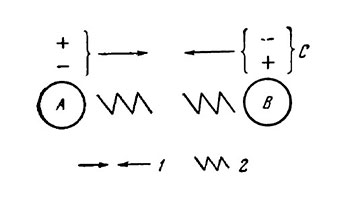
Рис. 2. Строение коллоидной частицы при образовании внутрикомплексного коацервата (Евреинова, 1954).
Л и В — коллоидные мицеллы; С — двойной слой:
1 — силы притяжения, 2 — силы отталкивания
Особый тип коацерватов, изучение которых имеет большое значение для понимания физико-химического состояния протоплазмы, представляют так называемые внутрикомплексные коацерваты (Опарин и Евреинова, 1954). Они образуются при адсорбции противоположно заряженных ионов на коллоидных мицеллах; последние как бы покрыты двойным слоем ионов. Многочисленные опыты показали, что коацерваты имеют ряд общих свойств с протоплазмой и что образование организованных структур в клетке также обусловлено процессом коацервации (рис. 3) (Boij, Bungenberg de Jong, В частности, соединение белковых растворов с нуклеиновыми кислотами дает внутрикомплексные коацерваты, в которых на белковых молекулах адсорбируются молекулы нуклеиновых кислот. Исследования коацерватных мицеллей дают возможность предположить, что коацерваты протоплазмы представлены коацерватами сложного типа, а коацерваты ядра — внутрикомплексными.
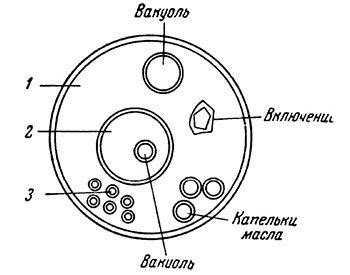
Рис. 3. Модель клетки по Бунгенберг дс Ионгу (Евреинова, 1954): 1, 2, 3 — коацерваты
Имеется еще одно существенное свойство коллоидных растворов, которое сближает их с физико-химическим состоянием протоплазмы. Это свойство связано со способностью коллоидных растворов находиться в двух состояниях — золя и геля. Золь — это жидкое состояние коллоида, в котором он обладает вязкостью (как и все жидкости); гель — это твердое (или полутвердое) состояние коллоида, в котором он проявляет себя как твердое тело, т. е. обладает эластичностью, стремлением сохранить свою форму, растяжимостью и т. д. Отличительной особенностью коллоидов является способность переходить от состояния золя к гелю, и наоборот (так называемый «золь-гель переход»). Золь-гель переход в коллоидных системах можно вызвать различными способами: изменением температуры или концентрации водородных ионов, прибавлением электролитов, механическим воздействием. Превращение геля в золь при механическом воздействии называется Это явление имеет большое значение для понимания физико-химического состояния протоплазмы. Например, многочисленные функционально-морфологические изменения протоплазмы можно рассматривать с точки зрения поведения тиксотропного геля.
Такими свойствами коллоидов, как аномальная вязкость (зависимость вязкости от давления), двойное лучепреломление, тиксотропные свойства, обладает и протоплазма живых клеток (Кедровский, 1946). Так, отчетливо было показано двойное лучепреломление митотического веретена, оболочки ядра, митохондрий, отмечено наличие аномальной вязкости у протоплазмы харовых водорослей. Тиксотропные свойства протоплазмы были обнаружены при исследовании разного рода течений протоплазмы, прямыми опытами с помощью микроманипуляции, определением вязкости (Гельбрунн, 1957).
Таким образом, протоплазму можно рассматривать как полифазную коллоидную систему, состоящую из высокомолекулярных соединений, диспергированных в водной среде.
Физико-химические представления о структурных свойствах протоплазмы должны быть тесно увязаны с рассмотрением надмолекулярных субклеточных структур. Электронно-микроскопические исследования показали наличие в протоплазме клетки различных нитевидных, пузырьковых, трубчатых, мембранных и другого рода структурных образований. Поэтому биологические процессы, протекающие в клетке, определяются не только ее коллоидно-физическими свойствами, по и физико-химическими явлениями, происходящими на многочисленных поверхностях раздела, образуемых надмолекулярными структурами (Эльпинер, 1953).
Физико-химические свойства протоплазмы
Наши знания жизнедеятельности клетки основаны на данных о физико-химических свойствах ее компонентов. Получаемые экспериментальные результаты характеризуют физико-химическое состояние живой протоплазмы и описывают изменение ее оптических, электрических, термических и механических свойств в состоянии различной функциональной активности. Наибольшее внимание привлекают те физико-химические показатели, которые выражают структурные и электро-кинетические свойства живой клетки: вязкость, окислительно-восстановительный потенциал, концентрация водородных ионов, положение изоэлектрической точки. Рассмотрению многочисленных данных по физико-химическим свойствам клетки посвящены обстоятельные монографии (Макаров, 1948; Гейльбрунн, 1957; Hewitt, 1950; Small, 1955 и др.). Здесь мы остановимся кратко лишь на некоторых из характеристик и на том новом освещении в понимании этих характеристик, которое появилось в научной литературе последних лет.
Вязкость коллоидов протоплазмы характеризует их структурное состояние и действующие между ними силы. Изучение вязкости протоплазмы показало, что этот показатель тесно связан с физиологической активностью клетки. Биоколлоиды протоплазмы клетки, находящейся в покое, соответствуют состоянию золя, переход ее в активное состояние вызывает повышение вязкости протоплазмы вплоть до образования геля. Такая глубокая перестройка биоколлоидов протоплазмы объясняется наличием различных сил сцепления между ними (электростатических, водородных, стерических), сильно и слабо действующих, действующих на близких и далеких расстояниях (Штокмайер, 1961). Необычное поведение протоплазмы в виде жидкости ив то же время полутвердого тела рассматривается как проявление структурной вязкости. Структурная вязкость протоплазмы связана со способностью протоплазменных мицелл образовывать нестойкие временные связи. Характерно, что указанное свойство принадлежит высокопоимерным соединениям — биологическим макромолекулам (белкам, нуклеиновым кислотам). Многообразные изменения вязкости, наблюдаемые в живых клетках, связаны со специфической способностью макромолекул к локальному изменению их структуры и обратимым изменениям их биологических функций. При изучении вязкости протоплазмы в настоящее время учитывается и то, что составляющие протоплазму биологические макромолекулы представляют собой полиэлектролиты, способные менять свою конфигурацию, сокращаться, растягиваться при изменении концентрации водородных ионов в среде (Волькенштейн, 1958).
Изоэлектрическая точка (ИЭТ) биоколлоидов является основной электроколлоидно-химической характеристикой клеточных компонентов. С физико-химической точки зрения она определяет равновесное состояние противоположно заряженных групп амфолита.
При цитохимическом анализе ИЭТ определяется с помощью окрашивания клеток растворами основных и кислых красителей различного pH.
Изучение сложных взаимоотношений белков, нуклеиновых кислот и липидов в клетке позволило по-новому оценить значение ИЭТ. Изменения связывания красителей в зависимости от pH дают возможность отличать различные белки (Singer, 1957), в то же время ИЭТ может характеризовать нуклеопротеидный комплекс, поскольку связывание основных красителей пропорционально количеству и степени диссоциации фосфорнокислых групп (Шабадаш, 1956). Кроме того, исследования показали, что ИЭТ, характеризующая электроколлоидальные свойства цитоплазмы, обусловлена по крайней мере тремя факторами — наличием свободной рибонуклеиновой кислоты, относительным содержанием рибонуклеопротеидов и составом белкового комплекса, а также его структурным состоянием (Конарев, 1958).
Окислительно-восстановительный потенциал имеет большое значение для жизнедеятельности клетки, так как все основные обменные процессы связаны с различными реакциями окисления и восстановления. Окислительно-восстановительный потенциал зависит от парциального давления кислорода, концентрации энзимов и метаболитов в клетке. Последовательность окислительно-восстановительных реакций в клетке определяется теми потенциалами, которые устанавливаются в тех или иных ферментных системах или субстратах.