
Исследования клетки и ее органоидов начаты с момента ее открытия. Из-за очень малых размеров субклеточных структур при исследовании их приходилось преодолевать огромные теоретические и методические трудности. Основным орудием исследования клетки был микроскоп, и успехи в микроскопической технике определяли прогресс науки о клетке. Когда была достигнута предельная разрешающая способность микроскопа, были сформулированы и основные представления о структурной организации клетки.
В меристематической клетке оказались хорошо видимыми клеточная оболочка, ядро и цитоплазма, а во взрослой клетке — вакуоль, занимающая значительную часть ее внутреннего объема.
В ядре, имеющем обычно шаровидную форму, хорошо видны одно или несколько ядрышек. При делении ядра легко обнаруживаются такие структурные образования, как хромосомы и веретено. В цитоплазме при соответствующем увеличении микроскопа можно видеть следующие субклеточные структуры: 1) пластиды; 2) митохондрии — сферические, палочковидные или нитевидные органоиды; 3) сферические секреторные гранулы, отличаемые от митохондрий только различным отношением к прижизненным красителям; 4) капельки жира и зерна крахмала, которые легко отличить от органоидов клетки по их оптическим свойствам и реакциям на окрашивание; 5) аппарат Гольджи (в животных клетках), представляющий собой сеточку из переплетенных нитей, или же имеющий вид гранул, часто серповидной формы, расположенных в цитоплазме группой или отдельными гранулами.
Несмотря на огромное разнообразие клеток тела растений, все они оказались построенными по единому плану, всем им свойствен определенный набор органоидов. Кроме того, на уровне субклеточных структур стираются различия между растительной и животной клеткой. В структуре и функции таких органоидов, как ядра, митохондрии и рибосомы, принципиальных отличий между животной и растительной клеткой пока не обнаружено.
В течение длительного времени, широко используя методы гистохимии, исследователи пытались выяснить химический состав видимых органоидов клетки и, главное, подойти к изучению их функции.
Совершенствование микроскопической техники (интерференционная микроскопия, ультрамикроскопия, поляризационно-оптические исследования, рентгеноскопия) не принесло решающих сдвигов в расшифровке структуры цитоплазмы, в познании формы и функции видимых органоидов клетки.
Обнадеживающие результаты по изучению химического состава клетки и ее органоидов были получены при применении флуоресцентной и ультрафиолетовой микроскопии, позволивших несколько шире интерпретировать функцию отдельных органоидов клетки.
Новый, исключительно продуктивный этап в исследовании структурной организации клетки начался с момента изобретения электронного микроскопа и разработки способа выделения органоидов клетки в количествах, достаточных для проведения почти всех обычных биохимических исследований.
Метод фракционного, или дифференциального, центрифугирования, использование изотопов в сочетании с электронным микроскопом позволили обнаружить в цитоплазме новые структурные образования и развернуть широким фронтом изучение функции отдельных органоидов. Если в недалеком прошлом изучением органоидов клетки занимались главным образом цитологи, то новые методы позволили включиться в эти исследования физиологам, биохимикам и биофизикам. В последнее десятилетие в познании клетки было сделано значительно больше, чем за предыдущие сто лет.
Для общей характеристики современного представления о структуре растительной клетки, основанного на данных электронной микроскопии, мы приведем ее схематическое изображение (рис. 4), взятое нами из доклада Ж. Дюсе на V Международном биохимическом конгрессе. В целях более целеустремленного рассмотрения значимости отдельных структурированных субклеточных систем в обмене веществ. В клетке Д. Е. Грин (1961) разделил их на три большие функциональные группы: 1) преобразующие машины (например, митохондрии и хлоропласты), катализирующие трансформацию энергии; 2) реплицирующие машины (например, рибосомы), катализирующие репликацию белков и других биополимеров; 3) интегрированные метаболические системы (например, клеточные гранулы), участвующие в синтезе жирных кислот и холестерина и катализирующие мультиэнзимные синтетические процессы. Несомненно, что такая группировка позволила дать Грину развернутую картину структуры и организации митохондрий как преобразующих энергию машин.
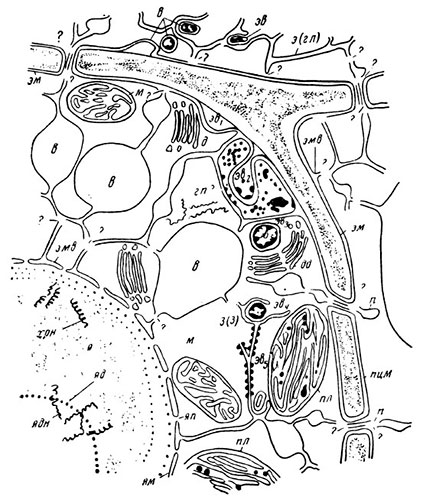
Рис. 4. Ультраструктура цитоплазмы растительных клеток (схема):
э (з) — эргастоплазма с зернистой поверхностью (с гранулами Палада); э (гл) — эргастоплазма с гладкой поверхностью (без гранул Палада): вместе они образуют сеть (эндоплазматический ретикулум), которая переходит в ядерную мембрану ям (эргастоплазматической природы); эм — экстоплазматическая мембрана, впячивания (эмз) и выросты (змв) которой вместе с ядерной мембраной создают непрерывность эндоплазматического ретикулума; n — плазмадесмы, пронизывающие пекто-целлюлозную мембрану (пцм) с проходящими через них эргастоплазматическими трубочками (непрерывность цитоплазмы); м — митохондрии; пл — пластиды, рассеянные в основном веществе; д — диктиосомы; гг — гидрофобные гранулы, часто окруженные эргастоплазмой; эв — эргастоплазматические впячивания, представляющие собой, вероятно, поглощение жидкости, аналогичные пиноцитозу; эв1, эв2, эв3...... эв& — последовательные состояния поглощения капелек; дд — диктиосомы в состоянии деления; я — ядро; яд — ядрышко; хрн — хроматиновые нити (по Рису); ядн — ядрышковые нити (проблематично); цн — цитоплазма
тические нити (проблематично); яп — ядерные поры; гп — гранулы Палада (проблематично)
Однако при такой группировке неизвестно, куда же поместить основной органоид клетки — ядро. При этом как будто бы полностью исключается возможность трансформации энергии в других органоидах клетки, кроме митохондрий и хлоропластов. Обнаружение рибосом в митохондриях и ядре также нарушает принцип группировки, предложенный Грином.
Следовательно, она в какой-то степени помогает более целеустремленно рассмотреть ведущую, определяющую функцию отдельных органоидов клетки, но далеко не достаточна для понимания взаимодействия органоидов клетки и их роли в ее метаболизме.
Протоплазма
Основную роль среди всех компонентов протоплазмы играют белки. Эта главенствующая роль белков определяется теми исключительными свойствами, которыми они обладают: необычайной химической реактивностью, разнообразием форм, способностью к структурообразованию и специфической активностью. Высокая химическая реактивность белковых молекул находит свое отражение в том, что в протоплазме они всегда соединены в сложную комплексную систему с другими важными соединениями: нуклеиновыми кислотами, углеводами, липидами и т. д.
Эти комплексные системы под влиянием различных внешних воздействий способны ж обратимым изменениям формы, соединению и разрушению, ж изменению активности, выражающейся в усилении или уменьшении жизнедеятельности клетки в целом.
Представление о протоплазме как о сложной гетерогенной коллоидальной системе требовало четкого представления об основных компонентах этой системы: дисперсной фазе и дисперсионной среде. Прямые и косвенные методы исследования (микрургия, фазово-контрастная микроскопия, изучение двойного лучепреломления, структурной вязкости, рефракции и т. д.) подтверждали, с одной стороны, наличие в протоплазме субмикроскопических единиц нитевидной (фибриллярной) формы, а с другой стороны, показывали существование гранулярных (глобулярных) частиц. Вместе с тем, при электронномикроскопических исследованиях было обнаружено, что протоплазма может иметь разное строение: сетчатое, пластинчатое, фибриллярное, гранулярное. Ряд авторов считает, что такие различные проявления субмикроскопического строения протоплазмы зависят от основных структурных элементов протоплазмы — глобулярных и фибриллярных белков и их взаимных превращений. Наиболее полное выражение этот взгляд получил в предложенной Фрей-Висслингом теории соединения (Frey-Wyssling, 1957). Согласно этой теории, основной элементарной единицей субмикроскопической структуры протоплазмы являются глобулярные макромолекулы, состоящие из сильно извитых линейных полипептидных цепей. Такие глобулярные макромолекулы, соединенные в виде нитки бус, образуют фибриллы или пластины. Эти линейные образования способны к различным поперечным соединениям, в результате чего образуется более или менее твердая сеть («остов» протоплазмы) (Frey-Wyssling, 1953). Образование ансамбля таких упорядоченных соединений имеет не случайный характер, а следует определенным полям притяжения на поверхности сложных глобулярных единиц. По мнению Фрей-Висслинга, эти поля притяжения играют ту же роль, что и координационные числа в кристалло-образовании. Если макромолекулы являются сильными диполями, то такие поля притяжения находятся на их противоположных сторонах и в этом случае возможно образование цепей (и, следовательно, фибриллярных структур) из глобулярных единиц. Соединения из трех таких элементов образуют пористую пленку (рис. 5). Между соседними полипептидными цепями возможны 4 различных типа связи: 1) гетерополярные силы межмолекулярного воздействия, вызываемые диполями, вследствие гидратации «коллоидной системы; 2) гегерополярные валентные связи, чувствительные к изменению pH (эфирные, или солевые, связи); 3) гомеополярные валентные связи, зависящие от окислительно-восстановительного потенциала (дисульфидная связь); 4) термолабильные гомеополярные силы межмолекулярного взаимодействия (например, связь белковых молекул через метильные группы) (Фрей-Висслинг, 1950).
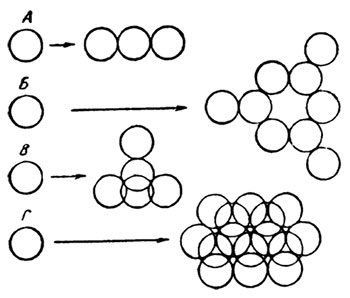
Рис. 5. Агрегация глобулярных макромолекул путем соединения.
На схеме показаны типы соединении глобулярных макромолекул:
А — цепочка; Б — пористая пленка; И — кристаллическая решетка; Г — сплошной слой
В связи с установлением того факта, что протоплазма состоит не из открытых полипептидных цепей, а из сильно спирализованных закрученных макромолекул, сейчас большое внимание уделяется не только внутримолекулярным силам, но и силам межмолекулярного взаимодействия (так называемым длиннодействующим силам (Вооф, 1961).
Электронномикроскопические исследования структуры цитоплазмы
Электронная микроскопия открыла необычайно сложный и разнообразный мир субмикроскопических структур протоплазмы. Детальные исследования этих субмикроскопических структур начались сравнительно недавно, однако уже сейчас электронно-микроскопические картины фиксированных клеток имеют большое постоянство своих основных черт и показывают четкие изменения субструктур при изменении функциональной активности клеток.
Приготовление биологического материала для исследования в электронном микроскопе требует особой обработки (фиксация, контрастирование, обезвоживание, заделывание в пластмассы или резины, просмотр среза в вакууме и т. д.). Естественно, что такая обработка вызывает различные изменения в исследуемом объекте и в основных веществах, составляющих его структуры, — белках, липоидах, нуклеиновых кислотах. Эти изменения должны строго контролироваться, поскольку при электронномикроскопическом исследовании объекта имеют дело с молекулярными масштабами, где определение размера той или иной структуры приобретает особо важное значение.
Как известно, протопласт растительной клетки включает в себя цитоплазму, ядро, вакуоль. Цитоплазма состоит из мезоплазмы, плазмалеммы и тонопласта.
Мезоплазма представляет собой центральную часть цитоплазмы, состоящую из основной цитоплазмы и цитоплазматических включений (пластиды, митохондрии, диктиосомы и т. д.).
Плазмалемма (эктопласт) и тонопласт представляют собой мембраны толщиной около 100 А, находящиеся соответственно на внешней и внутренней поверхностях мезоплазмы.
Основная цитоплазма (гиалоплазма) — это сложная гетерогенная система с целой серией субмикроскопических структур, доступных наблюдению только в электронном микроскопе. Все субмикроскопические структуры, находящиеся в основной цитоплазме, окружены непрерывной фазой, называемой цитоплазматической матрицей (или гиалоплазменной матрицей). В своем большинстве матрица состоит из макромолекулярной смеси, основой которой являются глобулярные белки, способные к обратимым глобулярно-фибриллярным изменениям. На электронномикроскопических фотографиях цитоплазматическая матрица видна в виде гомогенного или очень тонко зернистого вещества, имеющего низкую электронную плотность. Эта зернистость вследствие очень малого размера плохо поддается разрешению в электронном микроскопе. Зернистые образования могут наблюдаться в виде скоплений типа хлопьевидных агрегатов, но в этом случае они представляют собой артефакты, возникшие вследствие всей подготовительной обработки (Strugger, 1957).
В этой гомогенной матрице можно наблюдать различные четкие образования большой электронной плотности. Эти образования соединены в весьма сложную структурную организацию, пронизывающую всю мезоплазму. Одной из основных ультраструктур цитоплазмы является эндоплазматическая сеть (эндоплазматический ретикулум). Она представляет собой систему ограниченных мембранами пузырьков, трубочек, цистерн различного размера и формы, находящихся во всей основной цитоплазме (см. рис. 4). Было показано, что в растительных клетках эндоплазматическая вакуолярная сеть может быть связана с плазмалеммой и в то же время с ядерной оболочкой (Buvat, 1959). Это приводит исследователей к мысли о едином происхождении плазменной и ядерной оболочек и тесной связи всех структурных образований в функциональной деятельности клетки. На основе подобных исследований цитоплазма клетки рассматривается состоящей из двух структурных фаз, разделенных непрерывной системой мембран. Одна из этих фаз содержит цитоплазматическую матрицу и связана с ядром, а вторая, более или менее пустая, связана с внешним окружением (Mercer, 1960).
Другим важным компонентом цитоплазмы являются рибонуклеопротеидные частицы, или рибосомы. Рибосомы представляют собой частицы высокой электронной плотности диаметром около 150 А, которые либо находятся свободно в цитоплазме, либо связаны с внешней стороной мембран эндоплазматической сети. При дифференциальном ультрацентрифугировании рибосомы и мембраны эндоплазматической сети выделяются в виде особой фракции микросом, свойства которой подробно рассмотрены ниже (стр. 39). Функциональное значение рибосом окончательно еще не выяснено, но показана их тесная связь с синтезом белка в клетке и общими ростовыми процессами (Setterfeld, 1961).
В связи с рассматриваемым и субмикроскопическим и структурами заслуживает внимания анализ электронномикроскопических изображений клеток меристемы кончика корня луковицы (Allium сера) ио Штруггеру (Strugger, 1957). В гомогенной цитоплазматической матрице можно наблюдать глобулярные образования большой электронной плотности. Эти образования часто располагаются в виде одинарных или двойных рядов, соединенных поперечными связями, либо образующих небольшие нити, серповидные кружки и т. д. Анализируя электронномикроскопические фотографии, Шгруггер приходит к заключению, что в цитоплазме существуют субмикроскопические спирально закрученные нити (рис. 6).
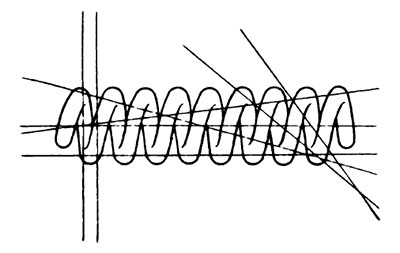
Рис. 6. Схематическое изображение спирально закрученной цитонемы. Прямые линии показывают возможные плоскости срезов цитонемы
Подобные спирализованньпе нити образуют основную массу субмикроскопических структурных элементов цитоплазмы, так называемую «цитонему» (рис. 7). Диаметр этих нитей 150-180 А, а диаметр образуемых ими спиралей в среднем 430 А; о длине их трудно сейчас сказать поскольку исследовались ультратонкие срезы, но максимальная длина равняется 3135 А. Таким образом, все видимые на ультратонких срезах глобулярные элементы представляют собой срезы основных спирально закрученных нитей, сделанных в различных плоскостях (рис. 8).

Рис. 7. Схематическое масштабное изображение субмикроскопического строения цитоплазмы меристематических клеток кончика корня луковицы:
1 — вакуоли, 2 — митохондрия, 3 — сферозома, 4 — диктиосома, 5 — звездообразное тело, 6 — двойные ламелли, 7 — цитонема
Эти заключения Штруггера совпадают со взглядами других исследователей, указывающих, что принцип спиральной структуры является самым характерным признаком субмикроскопической морфологии. Наглядным доказательством этого является обнаружение спиральной структуры α- и β-белков, нуклеиновых кислот и даже вирусных частиц (Frey-Wisling, 1957). Интересные подсчеты были сделаны при рассмотрении протоплазмы растительных клеток (Brown, 1960). Если принять среднее содержание белка в клетке равным 2×10-9 г, то площадь, которую займет это количество белка в виде монослоя, составит 400 000 мк2. Если принять среднюю площадь клетки равной 20 000 мк2, то, следовательно, в пределах клеточной стенки могут расположиться около 20 последовательных слоев белка. Даже если увеличить это число в 2,5 раза (50 слоев), то трудно представить, как они могут быть построены в сложный структурный комплекс эндоплазматической сети. Вместе с тем, хотя эндоплазматическая сеть хорошо наблюдается на ультратонких срезах и, как свидетельствуют электронные микрофотографии, весьма чувствительна к деформации, изменению условий питания, температуры, нет доказательств ее высокой лабильности. Если считать эндоплазматическую сеть твердой структурой, то трудно объяснить изменение таких физических характеристик, как структурная вязкость, переход золя в гель и т. д. Все это указывает на большую сложность, с которой сталкиваются цитофизиологи при исследовании клетки на ультраструктурном уровне, и свидетельствует о той большой работе, которую предстоит провести, чтобы воссоздать правильную и полную картину ультраструктурной организации клетки, тесно увязанную с современными физико-химическими данными о живой протоплазме.

Рис. 8. Схематическое изображение некоторых возможных типов плоских срезов цитонемы различного наклона и положения (Struger, 1.957)
Органоиды растительной клетки
Сохраняя историческую последовательность в изучении органоидов клетки, мы рассмотрим сначала структуру и функцию ядра, затем митохондрий, рибосом, аппарата Гольджи и некоторых других ее органоидов.