

В этой главе рассмотрены основные формы микроэлементов в растениях и механизмы их поглощение корнями. Описаны виды взаимодействий между химическими элементами на стадии их поглощения растением. Кроме того, содержится информация о физиологических и биохимических функциях микроэлементов в растительных тканях. Проанализированы закономерности распределения микроэлементов в различных частях растения. Указаны симптомы дефицита микроэлементов у растений и условия его возникновения.
Поглощение
В общих чертах механизмы поглощения растением микроэлементов существенно не отличаются от установленных для макроэлементов. В настоящее время получила распространение концепция мультифазного поглощения микроэлементов. Количество фаз поглощения и величины кинетических констант (Км и Vmax) значительно варьируют даже для одного и того же иона в зависимости от особенностей генотипа, возраста растений, диапазона концентраций, продолжительности эксперимента, взаимодействия с другими ионами (рис. 3.1).
Выделяют следующие основные фазы (табл. 3.1) в кинетике поглощения микроэлементов растениями: 1) фаза обменной адсорбции (продолжительность до 20 мин); 2) фаза насыщения свободного пространства (1-3 ч); 3) и 4) — фазы активного поглощения (до 6 ч и более).

Рис. 3.1. Поглощение цинка (1,3) и марганца (2,4) интактными корнями (1,2) и изолированными клеточными оболочками (3,4) кукурузы [по: Рудакова и др., 1987]
В поглощении микроэлементов отмечают пассивный и активный компоненты, соотношение между ними может изменяться в зависимости от свойств и концентрации элемента, биологических особенностей и возраста культур. По-видимому, активный транспорт микроэлементов осуществляется с участием как ионных насосов, так и переносчиков, в частности белков с молекулярной массой около 60-70 кД. Активное поглощение многих ионов микроэлементов преобладает в диапазоне более низких концентраций (система I), а пассивное — более высоких (система II).
В результате последних исследований в области молекулярной биологии удалось выявить различные переносчики, участвующие в транспорте микроэлементов в клетку через плазмалемму. У растений стратегии I (гл. 6), поглощающих железо преимущественно в форме Fe (II), выявлен ген IRT 1, который кодирует образование белка-переносчика со значительно большей специфичностью к Fe (II), чем к Fe (III) и другим потенциальным субстратам. По этой причине повышенные концентрации в среде других физиологически активных ионов металлов (Cu+, Cu2+, Mn2+, Zn2+), кроме кадмия, существенно не ингибируют поглощение растениями Fe (II). Субстратом для этого транспортера может быть не только Fe (II), но и кадмий.
| Культура | Фаза | Vmax
мкмоль/г.ч |
Км
мкмоль |
Интервалы перехода,
мкмоль |
| Zn2+ | ||||
| Горох | 1 | 0,116 | 1,2 | 0,1-1,0 (скачок) |
| 2 | ||||
| 3 | 2,469 | 47,0 | 10—100 (скачок) | |
| 4 | 3,571 | 137,0 | 200-1000 | |
| Кукуруза | 1 | 0,025 | 0,20 | 0,1-1,0 (скачок) |
| 2 | 0,082 | 2,40 | 5 (скачок) | |
| 3 | 1,471 | 45,0 | 10-200 (скачок) | |
| 4 | 1,818 | 110,0 | 5000-1000 | |
| Мn2+ | ||||
| Горох | 1 | 0,069 | 0,34 | 0,1-1,0 (скачок) |
| 2 | 0,100 | 5,0 | 1-5 | |
| 3 | 2,857 | 12,5 | 10-200 (скачок) | |
| 4 | 3,704 | 454,0 | 500-1000 | |
| Кукуруза | 1 | 0,003 | 0,29 | 0,1-1,0 (скачок) |
| 2 | 0,028 | 2,50 | 1-10 | |
| 3 | 0,430 | 10,0 | 10-100 (скачок) | |
| 4 | 0,530 | 250,0 | 200-1000 | |
Для растений стратегии II (гл. 6) выделены гены ids 2 и ids 3, которые ответственны за кодирование синтеза мугеневой кислоты, участвующей в транспорте железа через плазмалемму злаков в форме комплекса Fe(III)—мугеневая кислота. До сих пор не ясно, в какой форме через плазмалемму в клетку поступает медь: в форме Cu (II) или Cu (I). Иными словами, необходимо ли для поглощения меди предварительное восстановление катиона Cu2+(основная доступная для растений форма меди в природных средах) до Cu+. Например, арабидопсис имеет Cu-переносчик СОРТ 1, сходный с Cu -переносчиком ctr 1р. Последний использует в качестве субстрата одновалентный катион меди. В восстановлении Cu (II) до Cu (1), возможно, участвует Fe (ІІІ)-редуктаза, локализованная на поверхности корней растений стратегии 1. Во всяком случае мутанты frd 1, которые не могут индуцировать Fe (ІІІ)-редуктазную активность в ответ на Fe-дефицит, не способны восстанавливать Cu (ІІ)-хелаты.
В качестве возможных переносчиков марганца через плазматические мембраны рассматривают белки, кодируемые генами Nramp (Natural resistance associated macrophage protein). Семейство таких белков, высоко консервативных в эволюции, обнаружено не только у растений, но и у дрожжей, бактерий, птиц, млекопитающих. Все члены этого семейства переносчиков участвуют в транспорте металлов, главным образом в форме двухвалентных катионов.
Несмотря на важную роль цинка в жизни растений, механизмы поглощения этого металла корнями изучены мало. Не известно, поступает цинк преимущественно по ионным каналам или с участием двухвалентных переносчиков. Кроме того, не известно, активный этот процесс или пассивный. Изолированы гены ZIP 1, ZIP 2 и ZIP 3, которые кодируют синтез транспортеров цинка. Экспрессия генов происходит в широком временном диапазоне, а также температурном и концентрационном. У белков не обнаружено поглотительной активности по отношению к железу.
Взаимодействие
Сбалансированность минерального питания необходима для нормальных роста и развития растений. В сложных по химическому составу почвенных растворах содержится множество элементов. Взаимодействие между ними оказывает существенное влияние на доступность элементов для растения, в частности на проявление их дефицита. Взаимодействие химических элементов, в том числе и микроэлементов, может быть антагонистическим или синергическим (рис. 3.2). Антагонизм проявляется, когда физиологический эффект суммы нескольких элементов меньше суммарного физиологического эффекта элементов, взятых в отдельности. О синергизме можно говорить тогда, когда физиологическое действие нескольких элементов больше, чем отдельно взятых. Дефицит того или иного элемента у растений возникает, когда взаимоотношения между элементами носят конкурентный, антагонистический характер.
Конкуренция между элементами может возникать уже на стадии их физико-химической адсорбции клеточными стенками. Этим структурам принадлежит ведущая роль в формировании доннановской фазы свободного пространства растительной клетки. Конкуренции ионов способствует ограниченное число адсорбционных мест и обратимость большинства сорбционных процессов.
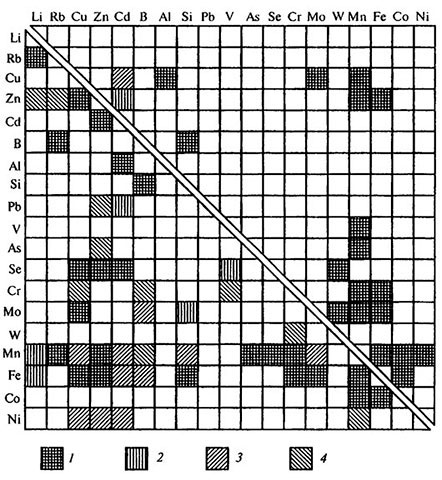
Рис. 3.2. Взаимодействие микроэлементов.
1 — антагонизм, 2 — синергизм, 3 — антагонизм и/или синергизм, 4 — возможный антагонизм [по: Кабата-Пендиас, Пендиас, 1989].
Связывание катионов микроэлементов клеточными стенками происходит с помощью ионизированных карбоксильных групп полисахаридов (пектиновых веществ, целлюлоз, гемицеллюлоз) и гликопротеинов, входящих в состав матрикса. Эти группы обеспечивают большое число отрицательных фиксированных зарядов в клеточной стенке. Поглощение катионов осуществляется пропорционально числу свободных карбоксильных групп в клеточных стенках. Микроэлементы могут связываться и карбоксильными группами белков. Однако эти группы сосредоточены главным образом в пектиновых веществах, во фракции которых концентрируются и микроэлементы. Удаление пектиновых веществ из клетки сопровождается резким снижением поглощения микроэлементов. Растения, содержащие в корнях много пектиновых веществ, как правило, поглощают и большие количества микроэлементов. Например, корни гороха адсорбируют значительно большие количества цинка и марганца, чем корни кукурузы. Эти различия объясняют большей (примерно на порядок) обогащенностью корней гороха пектиновыми веществами. С помощью специальных методик, позволяющих последовательно удалять из клеточных стенок различные функциональные группы, установлено, что разрушение только карбоксильных групп приводит к элиминированию поглощения ионов клеточными оболочками. Кроме карбоксильных групп в поглощении ионов принимают участие и другие функциональные группы, заряженные как отрицательно (гидроксильные, фосфатные), так и положительно (аминные). Последние придают клеточной стенке анионообменные свойства.
Прочность связи микроэлементов клеточными стенками сильно варьирует в зависимости от физико-химической природы ионов и их концентрации в среде. По устойчивости образуемых с клеточной стенкой связей двухвалентные катионы металлов-микроэлементов располагаются в ряду Cu2+ > Zn2+ > Fe2+ > Мn2+. С увеличением степени окисленности металлов их сродство к клеточным стенкам возрастает. Поглощение анионов микроэлементов (бор, молибден) из растворов с их низким содержанием сильно замедлено вследствие электростатического отталкивания одноименных зарядов анионов и клеточных стенок.
Наиболее прочные связи с клеточными оболочками формируются при низких концентрациях микроэлементов в среде. С повышением концентрации связи микроэлементов с клетками корней существенно ослабевают. Возможно, при низких концентрациях ионов в растворе сначала насыщаются сайты, дающие с катионами микроэлементов наиболее прочные связи, в том числе координационные. По мере насыщения первоначальных мест поглощения «включаются» другие функциональные группы, связи которых с катионами микроэлементов значительно слабее. Этот факт установлен для микроэлементов с разной комплексообразующей способностью (Zn, Мп, Li) на примере отделенных корней и изолированных клеточных оболочек гороха и кукурузы. При высоких концентрациях микроэлементов в среде на поверхности корней могут также образовываться нерастворимые соединения металлов.
Проявление конкуренции вероятнее всего между ионами, имеющими сходные свойства, механизмы поглощения и места связывания клеточными оболочками. Избирательность поглощения растениями ионов зависит от величины их заряда и радиуса, гидрационной и поляризующей способности, координационных свойств, коэффициентов диффузии. В результате поливалентные металлы сильнее экранируют отрицательный заряд клеточных стенок, чем одновалентные. Антагонизм между ионами, как правило, регистрируют при более высоких концентрациях в растворах, т. е. в диапазоне функционирования системы II, осуществляющей поглощение ионов в основном пассивным способом.
Взаимодействие элементов происходит не только на этапе поглощения корнем, но и при их транспорте и включении в метаболизм растения. Например, свинец подавляет как поглощение, так и передвижение железа, марганца и цинка в побег в результате нарушения связывания микроэлементов с комплексообразователями, участвующими в переносе ионов. Аналогичный факт установлен для магния по отношению к цинку. Конкуренция между металлами может проявляться в ферментных системах за места связывания с апобелком, что отражается на скорости биокаталитических реакций.
Конкуренция между ионами обычно сопровождается снижением их накопления в растениях (рис. 3.3; 3.4). Вследствие антагонизма железа с тяжелыми металлами у .растений появляется хлороз. Классическим примером антагонизма между ионами служат взаимоотношения железа и марганца.
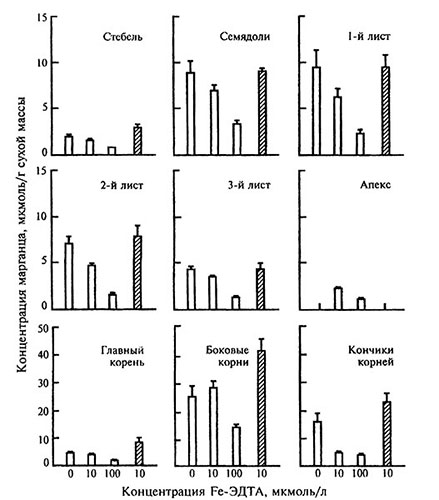
Рис. 3.3. Концентрация марганца в различных частях растений томата в зависимости от содержимого железа в питательном растворе [модифицировано по: Pich et al., 1994].
Светлые столбики — дикий вид, заштрихованые — мутант chloronerva, у которого симптомы хлороза проявляються даже при нормальном обеспеченности растений железом.
Из макроэлементов кальций, магний и фосфор выступают главными конкурентами микроэлементов в процессе поглощения растениями (табл. 3.2). Особая роль в регуляции избирательной специфичности оболочек растительной клетки принадлежит кальцию. Катионы кальция стабилизируют биологические мембраны и влияют на величину трансмембранного электрохимического потенциала. В случае недостатка ионов кальция в среде избирательность мембран при поглощении ионов понижена, что сопровождается интенсивным выделением из клетки относительно низкомолекулярных соединений. Для различных культур описана сильная конкуренция между марганцем, с одной стороны, и магнием и кальцием, с другой. Антагонизм ионов этих металлов обнаружен на стадии их поступления в клетку. В повышенных концентрациях кальций и магний могут ограничивать поступление марганца не только в корни, но и в побег. Напротив, с возрастанием концентрации марганца в среде подавляется поступление кальция и магния в растительные ткани, в том числе и в семена.
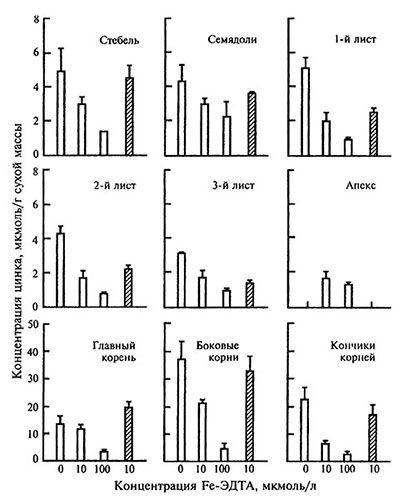
Рис. 3.4. Концентрация цинка в различных частях растений томата в зависимости от содержания железа в патительном растворе
Обозначение как на рисунке 3.3.
На поступление микроэлементов в растение и их аккумуляцию в растительных тканях сильное влияние оказывают тяжелые металлы. Многие из них (Cd, Pb и др.) не относятся к группе элементов, необходимых растениям, и в повышенных концентрациях фитотоксичны. Например, под влиянием кадмия значительно подавляется поглощение корнями многих микроэлементов (Fe, Мn, Zn), что сопровождается снижением их концентрации в растительных тканях. Вместе с тем токсичное действие тяжелого металла может приводить к относительному увеличению поступления микроэлементов в надземные органы по сравнению с корнем (например, марганца под воздействием увеличивающейся концентрации кадмия в питательной среде).
| Макроэлеметы | Микроэлементы и другие химические элементы | |
| Антагонизм | Синергизм | |
| Са | Аl, В, Ва, Be, Cd, Со, Cr,
Cs, Сn, F, Fe, Li, Mn, Ni, Pb, Sr, Zn |
Cu, Mn, Zn |
| Mg | AI, Be, Ba, Cr, Mn, F, Zn | AI, Zn |
| Р | AI, As, B, Be, Cd, Cr, Сn,
F, Fe, Hg, Mo, Mn, Ni, Pb, Rb, Se, Si, Sr, Zn |
AI, В, Cu, F, Fe, Mo, Mn, Zn |
| К | AI, B, Cd, Cr, F, Mo, Mn, Rb | — |
| S | As, Ba, Fe, Mo, Pb, Se | F, Fe |
| N | B, F, Сn | В, Cu, Fe, Mo |
Между анионами также наблюдается конкуренция, а значит, и ограничение селективности мест их связывания на плазматической мембране. Хорошо известны такие примеры конкуренции как взаимодействие сульфата и молибдата, сульфата и селената, фосфата и арсената. Повышенное содержание в почве сульфатов может существенно ограничивать поступление в растение молибдена.
Взаимодействие ионов не всегда приводит к одинаковым результатам. Например, описаны случаи не только антагонистического, но и синергического, а также индифферентного действия кальция на поглощение и транспорт ионов микро- и макроэлементов. Физиологическое проявление взаимодействия ионов зависит от многих факторов: биологических особенностей культур (в частности, их толерантности к действию тяжелых металлов), природы и концентрации в среде ионов, а также специфики органа или органеллы растения, где измеряли концентрацию элементов. Однозначное взаимодействие ионов регистрируется только при их определенных соотношениях и одинаковой длительности воздействия.