

В этой главе основное внимание сосредоточено на закономерностях распределения микроэлементов в семенах высших растений на разных стадиях их генеративного развития. Описаны пути транспорта микроэлементов в формирующихся семенах. Приведена информация о формах, в которых микроэлементы запасаются в семенах. Рассмотрены динамика распределения микроэлементов при прорастании семян и донорно-акцепторные отношения между органами семян и проростка. Определена роль отдельных частей семени в снабжении микроэлементами осевых органов зародыша.
4.1. Формирующиеся семена
Формирование семян высших растений начинается с момента оплодотворения и заканчивается образованием зрелых семян. Условно этот период Д. Бевли (1997) делит на три фазы. Во время фазы I происходит интенсивное деление клеток и формирование структур семени. После окончания интенсивного клеточного деления начинается фаза II. На этой стадии развития резервные вещества интенсивно аккумулируются в семенах, что сопровождается увеличением сухой массы последних. После накопления сухой массы начинается фаза III, в течение которой достигается физиологическая зрелость семян. Наступление этой фазы определяет, по существу, окончание продукционного процесса у культурных растений.
Особенности распределения микроэлементов по органам зрелого семени во многом зависят от транспорта микроэлементов в семенах в ходе их формирования. На стадии репродуктивного развития растений значительно снижается активность корней и соответственно поглощение питательных элементов из почвы. Поэтому при формировании семян возрастает роль ремобилизации минеральных веществ из других органов растений.
Под ремобилизацией понимают совокупность физиологических и биохимических процессов, направленных на вторичное использование минеральных элементов и запасных веществ формирующимися органами. Ремобилизация минеральных элементов включает следующие стадии: мобилизацию в индивидуальных клетках, ближний транспорт элементов по симпласту к флоэме, загрузку флоэмы и флоэмный транспорт. На стадии формирования семян интенсивность ремобилизации зависит от специфической потребности семян в конкретном элементе, минерального статуса вегетативных частей, мобильности элементов при движении по флоэме. У хлебных злаков свыше 90% общего фосфора в зерновках может обеспечиваться ремобилизацией этого элемента из вегетативных органов. Для азота эта величина составляет 50-90% в зависимости от условий культивирования растений. Высокой степенью ремобилизации характеризуются и некоторые микроэлементы. В плоды люпина поступает до 50% микроэлементов от их исходного количества в листьях. Скорость ремобилизации микроэлементов зависит также от минерального статуса листа. У пшеницы за весь период развития зерновки листья с высоким содержанием меди могут терять до 70% этого элемента, а с низким только 20%. Различия объясняют тем, что в листьях при дефиците микроэлементов велика доля их прочно связанных форм.
Мобильность химического элемента во флоэме играет важную роль в его транспорте от источника к емкости. Этот показатель зависит от генотипа и минерального статуса растений. Однако при передвижении по флоэме (опыты in vivo) наиболее высока мобильность хлора, которая сопоставима с мобильностью макроэлементов: калия, магния, фосфора, серы, азота. Низкая мобильность отмечена для марганца, а из макроэлементов — для кальция. Железо, цинк, медь, бор и молибден отличает средняя мобильность во флоэме. На репродуктивной стадии развития растений ремобилизация микроэлементов и кальция обычно выше, чем в период вегетативного роста. Формирующиеся семена и плоды индуцируют старение листьев, в которых изменяется гормональный баланс, играющий важную роль в ремобилизации. Старение листьев могут также усиливать затенение и дефицит питательных веществ (азота), что соответственно повышает степень ремобилизации микроэлементов из этих органов. Отметим, что до сих пор не установлен вклад флоэмного транспорта микроэлементов в их аккумуляцию в зерновке по сравнению с транспортом по ксилеме.
Основные исследования накопления микроэлементов в формирующихся семенах выполнены Д. Пирсоном и Р. Грахамом с коллегами (1994-1998) на пшенице. Марганец и цинк транспортируются к зерновкам различными путями. Цинк, транспортируемый по ксилеме, прежде чем поступить в зерновку пшеницы попадает во флоэму плодоножки и ости. С помощью ингибиторов мембранного транспорта было установлено, что марганец из ксилемы поглощается так называемыми переходными клетками, а затем по симпласту попадает в проводящую систему зерновки.
После проведения экспериментов с изотопами 56Мn и 65Zn удалось выяснить, что скорость поступления этих микроэлементов в зерновку на ранних стадиях генеративного развития пшеницы выше, чем на завершающих. Причем максимальное количество микроэлементов аккумулируется в наиболее интенсивно растущих в этот период тканях: в перикарпе, а также в эндосперме (рис. 4.1). Аккумуляция марганца и цинка в зародыше происходит на более поздних этапах развития зерновки.

Рис. 4.1. Аккумуляция 65Zn и 56Mn в тканях зерновки пшеницы в процессе их развития [по: Pearson et al., 1998]
Цинк и марганец, поступившие непосредственно в зерновку, в основном перемещаются по флоэме проводящей системы бороздки и накапливаются в тканях эндокарпа бороздки. В зоне бороздки располагается основная проводящая система плода злаков, состоящая из одного или нескольких пучков. При разрушении флоэмы бороздки над зародышем (введение внутрь проводящего пучка горячей иглы) резко снижается поступление марганца и цинка в эндокарп, а также марганца в зародыш (рис. 4.2).
В дорсальную часть перикарпа транспорт марганца и цинка может осуществляться по протофлоэме стенок завязи. Туда поступает менее 5% цинка и 15% марганца, транспортируемых в зерновку. В дорсальной области перикарпа располагаются три пучка протофлоэмы. Два из них локализованы на боковой стенке завязи и тянутся по всей длине зерновки. Третий пучок находится напротив складки на дорсальной стороне стенки завязи и простирается лишь на часть завязи. Эти пучки существуют до времени полусозревания зерновки или еще дольше, а затем постепенно отмирают с базипетального конца. В это время уменьшается поступление в перикарп цинка и марганца, особенно марганца. Возможно, марганец и цинк транспортируются в дорсальную часть перикарпа и через алейроновый слой зерновки: этот слой выделяет соединения, попадающие через эндокарп в экзокарп. Для подтверждения этого предположения необходимы дополнительные исследования.
Основной приток цинка в зародыш созревающей зерновки пшеницы осуществляется по флоэме пучка, начинающегося в плодоножке. Разрушение флоэмы бороздки в зоне выше зародыша практически не влияет на поступление микроэлемента в этот орган в ходе его развития (рис. 4.2).

Рис. 4.2. Аккумуляция 65Zn 54Mn в тканях зерновок пшеницы после разрущения флоэмы бороздки выше зоны зародыша [по: Pearson et al., 1998]
В то же время транспорт в зародыш марганца может проходить по флоэме как пучка плодоножки, так и бороздки. Поэтому разрушение последней существенно снижает поступление марганца в зародыш.
Интенсивное поступление микроэлементов в зерновку по срокам совпадает с их сильным оттоком из вегетативных органов. Пути транспорта в зерновке сахарозы и микроэлементов (Zn и Мn) отличаются. Сахароза преимущественно транспортируется по проводящей системе бороздки к эндосперму, где распределяется в радиальном направлении и запасается в виде крахмала. Микроэлементы, как уже упоминалось выше, поступают в эндосперм в значительно меньшем количестве.
На заключительных этапах марганец и цинк могут перераспределяться внутри зерновки (пшеница) с током воды. Ретранслокация из перикарпа отмечена, по крайней мере, для марганца. Аккумуляция цинка в зародыше и эндосперме происходит вследствие оттока этого металла из верхней и нижней цветковых чешуй, но не из перикарпа. Возможно, марганец не перемещается из вегетативных тканей вторичного колоска и накапливается в зерновке в результате транспорта из фондов в плодоножке и ости. По мере созревания зерновки клетки перикарпа разрушаются и большинство его ассимилятов попадает в эндосперм. Несмотря на возможность ремобилизации микроэлементов из перикарпа на завершающей стадии развития, в зрелых зерновках этот орган сохраняет значительные количества микроэлементов (см. 4.2). Поэтому использование технологий помола, в которых перикарп изымается (лущение), приводит к сильному обеднению микроэлементного состава муки.
4.2. Зрелые семена
4.2.1. Общее содержание микроэлементов
Уровень содержания микроэлементов в семенах и вариабельность микроэлементного состава растений в основном определяются генотипическими особенностями культур, возрастом растений и экологическими факторами.
Принято считать, что у высших растений элементный состав репродуктивных органов отличается большим постоянством, чем таковой вегетативных. Относительный консерватизм генеративных органов обусловлен более строгим генетическим контролем. В условиях острого дефицита минеральных элементов формирование генеративных органов заторможено или вовсе не происходит. Избыточному накоплению элементов в семенах и плодах препятствуют барьерные функции вегетативных органов, например концентрирование металлов в апопласте корней.
Вместе с тем содержание микроэлементов в семенах растений варьирует в довольно широком диапазоне (табл. 4.1; 4.2), что обусловлено генетическими и экологическими факторами. У злаков высокие концентрации железа обнаружены в зерновках ячменя и риса, а марганца в зерновках овса (табл. 4.1).
| Элемент | Концентрация, мг/кг сухой массы | ||
| Средняя | Минимальная | Максимальная | |
| Пшеница | |||
| Fe | 37,5 ± 3,1 | 20,7 ± 2,7 | 64,5 ± 9,6 |
| Мn | 38,6 ± 3,4 | 21,3 ± 4,2 | 71,1 ± 8,4 |
| Сu | 4,99 ± 0,29 | 2,78 ± 0,34 | 8,05 ± 1,19 |
| Zn | 30,1 ± 2,1 | 19,8 ± 1,9 | 47,4 ± 4,9 |
| Со | 0,104 ± 0,032 | 0,079 ± 0,021 | 0,252 ± 0,076 |
| Мо | 0,382 ± 0,063 | 0,197 ± 0,038 | 0,872 ± 0,165 |
| В | 1,44 ± 0,32 | 0,78 ± 0,36 | 1,62 ± 0,53 |
| Ячмень | |||
| Fe | 74,8 ± 13,7 | 35,1 ± 5,8 | 120,8 ± 20,9 |
| Мn | 30,3 ± 4,8 | 15,5 ± 2,3 | 50,8 ± 12,7 |
| Сu | 6,03 ± 0,79 | 3,61 ± 0,58 | 9,80 ± 2,07 |
| Zn | 30,3 ± 2,7 | 19,9 ± 3,0 | 37,7 ± 3,7 |
| Со | 0,14 ± 0,09 | 0,07 ± 0,04 | 0,21 ± 0,13 |
| Мо | 0,44 ± 0,14 | 0,18 ± 0,04 | 0,66 ± 0,17 |
| В | 2,74 ± 0,60 | 1,72 ± 0,59 | 2,87 ± 1,06 |
| Овес | |||
| Fe | 48,9 ± 4,6 | 37,3 ± 4,8 | 75,1 ± 10,0 |
| Мn | 54,8 ± 10,0 | 28,3 ± 7,8 | 117,9 ± 30,5 |
| Сu | 6,15 ± 0,69 | 3,85 ± 0,73 | 9,11 ± 1,19 |
| Zn | 26,7 ± 6,8 | 18,1 ± 5,1 | 48,6 ± 17,2 |
| Со | 0,080 ± 0,001 | 0,027 ± 0,007 | 0,113 ± 0,028 |
| Мо | 0,48 ± 0,10 | 0,23 ± 0,06 | 0,73 ± 0,13 |
| В | 2,24 ± 0,51 | 1,06 ± 0,53 | 2,41 ± 0,80 |
| Кукуруза | |||
| Fe | 28,1 ± 4,3 | 18,2 ± 1,7 | 30,4 ± 2,8 |
| Мn | 9,2 ± 1,9 | 6,1 ± 1,4 | 9,5 ± 1,5 |
| Сu | 2,91 ± 0,47 | 1,94 ± 0,53 | 3,44 ± 0,29 |
| Zn | 28,3 ± 3,0 | 23,4 ± 2,4 | 40,3 ± 4,8 |
| Со | 1,92 | 1,82 | 2,02 |
| Мо | 0,142 ± 0,044 | 0,070 ± 0,031 | 0,403 ± 0,208 |
| В | 2,93 ± 0,07 | 2,50 ± 0,50 | 3,50 ± 0,50 |
| Рис | |||
| Fe | 58,4 ± 19,4 | 38,0 ± 10,3 | 90,6 ± 38,5 |
| Мn | 35,1 ± 10,9 | 18,4 ± 5,4 | 79,1 ± 29,6 |
| Сu | 5,95 ± 1,09 | 3,69 ± 0,76 | 7,92 ± 0,86 |
| Zn | 25,5 ± 3,0 | 17,9 ± 2,3 | 35,5 ± 4,3 |
| Мо | 0,74 | 0,24 | 3,23 |
Среди бобовых содержание бора, железа, меди и цинка в семенах сои обычно выше, чем в семенах гороха и фасоли (табл. 4.2). Самые высокие концентрации кобальта установлены в семенах фасоли. Среднее содержание молибдена в семенах бобовых уменьшается в ряду: горох > соя > фасоль. В целом в семенах бобовых содержание микроэлементов выше, чем в зерновках злаков. Наиболее сильные различия между указанными группами растений установлены по содержанию в семенах молибдена и бора. Не случайно поэтому бобовые культуры относят к растениям — концентраторам микроэлементов. Выявленные закономерности следует рассматривать как ориентировочные, поскольку они базируются на результатах усреднения многочисленных данных, полученных с использованием различных методических подходов.
Содержание микроэлементов в семенах может варьировать у растений не только разных видов, но и в пределах одного вида. Так, микроэлементный состав семян существенно варьирует у пшеницы, кукурузы, сои и других культур различных сортов.
| Элемент | Концентрация, мг/кг сухой массы | ||
| Средняя | Минимальная | Максимальная | |
| Горох | |||
| Fe | 56,4 ± 5,0 | 36,6 ± 6,3 | 75,2 ± 8,5 |
| Мn | 20,9 ± 2,6 | 10,5 ± 1,1 | 23,7 ± 1,8 |
| Сu | 10,2 ± 1,0 | 6,2 ± 0,7 | 15,7 ± 1,6 |
| Zn | 36,4 ± 2,4 | 28,6 ± 4,3 | 55,6 ± 5,6 |
| Со | 0,31 ± 0,08 | 0,23 ± 0,06 | 0,63 ± 0,22 |
| Мо | 5,4 ± 0,8 | 1,35 ± 0,41 | 14,53 ± 5,39 |
| В | 7,00 ± 0,78 | 6,48 ± 0,48 | 12,78 ± 2,49 |
| Соя | |||
| Fe | 131,9 ± 23,5 | 86,0 ± 11,4 | 178,0 ± 40,1 |
| Мn | 31,2 ± 2,7 | 19,5 ± 2,3 | 45,8 ± 5,4 |
| Сu | 14,8 ± 1,6 | 9,4 ± 1,8 | 19,3 ± 2,2 |
| Zn | 51,3 ± 3,7 | 33,7 ± 3,2 | 67,6 ± 8,9 |
| Со | 0,37 ± 0,08 | 0,13 | 0,46 |
| Мо | 3,89 ± 1,08 | 1,04 ± 0,53 | 13,13 ± 3,00 |
| В | 24,4 ± 2,7 | 23,0 ± 3,8 | 40,3 ± 6,3 |
| Фасоль | |||
| Fe | 61,7 ± 10,5 | 61,1 ± 20,5 | 98,1 ± 24,7 |
| Мn | 17,5 ± 3,2 | 6,0 | 87,0 ± 67,0 |
| Сu | 8,5 ± 0,6 | 3,6 ± 1,3 | 9,9 ± 1,8 |
| Zn | 32,4 ± 4,4 | 29,4 ± 6,2 | 46,3 ± 6,8 |
| Со | 0,73 ± 0,54 | 0,45 ± 0,32 | 9,17 ± 4,47 |
| Мо | 2,42 ± 0,54 | 0,27 ± 0,14 | 8,16 ± 1,29 |
| В | 15,8 ± 4,7 | 14,7 ± 1,3 | 25,3 ± 4,7 |
Уровень накопления и состав микроэлементов в семенах в значительной степени зависят и от почвенно-климатических факторов. В зоне тайги и подтайги в зерновках пшеницы, ячменя, ржи и овса содержится больше марганца, меньше меди и молибдена, по сравнению с лесостепью и степью. В зоне тайги и подтайги обнаружено и более низкое содержание меди в семенах гороха.
Встречаются сведения о довольно сильных колебаниях в микроэлементом составе семян растений в разные годы. Реакция почв и содержание в них карбонатов являются важными экологическими факторами, влияющими на состав семян растений (табл. 4.3). У растений, выращенных на карбонатных почвах, более низкая концентрация в семенах кобальта и марганца, а также цинка.
| Элемент | Концентрация, мг/кг сухой массы | ||
| Карбонатные почвы, pH 7,5-8,5 | Кислые почвы, pH 4,5-6,0 | ||
| Калий | 229 | 228 | |
| Рубидий | 0,14 | 0,32 | |
| Магний | 95 | 104 | |
| Кальций | 246 | 199 | |
| Марганец | 0,9 | 1,4 | |
| Железо | 1,3 | 1,3 | |
| Кобальт | 0,0021 | 0,0045 | |
| Медь | 0,17 | 0,19 | |
| Цинк | 0,53 | 0,65 | |
| Молибден | 0,016 | 0,013 | |
| Бор | 3,4 | 3,3 | |
| Фосфор | 134 | 152 | |
| Сера | 77 | 76 | |
В условиях агроценозов содержание микроэлементов в семенах растений зависит от вида и способов применения макро- и микроудобрений. Спектр эффектов, вызываемых макроудобрениями, весьма широк. Под влиянием внесения в почву полного удобрения (N, Р, К) увеличивается содержание в зерновках озимой пшеницы марганца и молибдена. Под влиянием фосфорных удобрений снижается содержание в семенах разных видов растений железа и цинка. Во многих случаях увеличение содержания микроэлемента в удобрениях сопровождается его накоплением в семенах. Масштабы накопления зависят от способа применения микроудобрений. Так, наиболее существенное запасание молибдена в семенах люпина происходит при некорневой подкормке этим элементом, а не при предпосевной обработке семян.
На микроэлементный состав семян влияют взаимоотношения: синергизм или антагонизм, элементов в системе почва—растение (см. 3.1). Например, содержание марганца в зерновках кукурузы при внесении в почву молибденовых удобрений увеличивается (синергизм), а содержание железа в зерновках риса и кукурузы при внесении цинковых удобрений уменьшается (антагонизм). Фосфор негативно влияет на поступление в семена цинка и марганца, а сера на поступление молибдена. Конкуренция микроэлементов происходит не только на стадии их поглощения корнем, но и при транспорте в растении. Микроэлементы, нанесенные на поверхность вегетативных органов (некорневые подкормки), влияют на элементный состав семян. Так, при опрыскивании растений ячменя соединениями меди в зерновках достоверно снижается концентрация марганца. В свою очередь, поступление в зерновки меди ингибируют кобальтовые удобрения.
Несмотря на то что экологические факторы оказывают существенное влияние на элементный состав семян, растительный организм может в какой-то мере поддерживать необходимый уровень содержания минеральных элементов в семенах. Некоторые элементы, находящиеся в среде в малоподвижной форме, преимущественно концентрируются в семенах. Такой вывод был сделан на основании анализа отношения концентраций микроэлементов в семенах и листьях растений, выращенных на карбонатных и кислых почвах (рис. 4. 3). В условиях дефицита (карбонатные почвы) концентрация марганца, железа, бора в листьях уменьшалась в большей степени, чем в семенах. Как известно, подвижность указанных элементов в почве очень сильно снижается с повышением pH среды. В семенах концентрация микроэлементов может стабилизироваться в результате их перераспределения между вегетативными и генеративными органами.
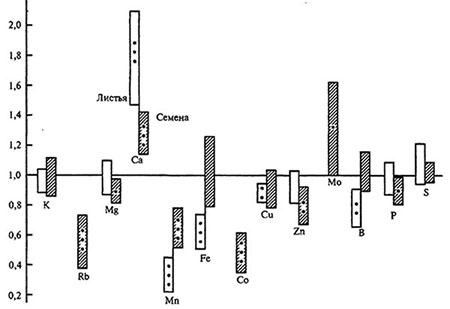
Рис. 4.3. Средние отношения (корбонатная почва: кислая почва) концентраций элементов в листьях и семенах растений 35 видов семейств Asteracea, Apiacea, Papilionacae, Caryophyllaceae, Rosaceae, Polygonaceae [по: Tyler, Zohlen, 1988].
Достоверност различий: *p < 0,05, **p < 0,01, ***p < 0,001
4.2.2. Распределение
Относительно микроэлементов, в отличие от органических веществ (белков, углеводов, липидов) и макроэлементов (N, Р, К, Са, Mg), этот вопрос слабо изучен. Основные сведения получены на примере зерновок злаков. В зрелых зерновках злаков наиболее высокие концентрации микроэлементов (расчет на единицу сухой массы), как правило, регистрируют в тканях семенных покровов и зародыша, а наиболее низкие в тканях эндосперма (табл. 4.4). Высокие концентрации ряда микроэлементов в зародыше отмечены и для некоторых видов бобовых растений. Вместе с тем у злаков основным запасающим органом многих минеральных элементов, в том числе и микроэлементов, служит эндосперм — самый крупный орган зерновки злаков, занимающий около 83% объема всего плода. У кукурузы запасы цинка в эндосперме и зародыше сопоставимы, что связано с высокой концентрацией этого элемента в тканях зародыша
| Микроэлемент | Концентрация, мг/кг сухой массы | ||
| Зародыш | Эндосперм | Семенные покровы | |
| Fe | 146,1 ± 9,8 | 46,6 ± 4,9 | 136,3 ± 10,6 |
| Мn | 25,8 ± 3,2 | 13,7 ± 2,2 | 30,2 ± 4,4 |
| Zn | 55,6 ± 7,3 | 9,1 ± 1,2 | 41,6 ± 1,3 |
Прим. Концентрация микроэлементов в частях зерновок определена рентгенофлуоресцентным методом через 24 ч после замачивания зерновок в воде. Приведены средние для растений семи генотипов, выращенных в одинаковых условиях. Ошибка ± t sx.
Прим. Концентрации определены с помощью протонного микроанализа. Ошибка ± 1 sd.
При использовании сканирующего электронного микроскопа в комбинации с рентгеновским микроанализом была получена информация о детальном распределении микроэлементов в пределах отдельных органов зрелой зерновки (табл. 4.5). Так, для пшеницы наибольшая концентрация марганца отмечена в зародышевом корешке (рис. 4.4), а наименьшая в колеоризе. Концентрация цинка выше в щитке, ниже в листовом примордии. Самая высокая концентрация железа установлена в сосудистом пучке и щитке, а самая низкая в колеоризе и листовых примордиях.
| Часть зерновки | Концентрация, мг/кг сухой массы | |||
| Мn | Fe | Сu | Zn | |
| Эндосперм | (3 ± 1)×10 | <4×10 | <8 | (4 ± 1)×10 |
| Зародышевый корешок | (1,4 ± 0,2)×103 | (2,1 ± 0,8)×102 | (2 ± 1)×10 | (5,5 ± 0,9)×102 |
| Проводящий пучок | (6,0 ± 0,9)×102 | (5 ± 2)×10 | (4,0 ± 0,8)×10 | (1,2 ± 0,2)×102 |
| Щиток | (5,7 ± 1)×102 | (5,7 ± 1,0)×102 | (8 ± 2)×10 | (6 ± 1)×102 |
| Колеориза | (2,0 ± 0,3)×102 | (1,5 ± 0,4)×102 | <4×10 | (4,2 ± 0,7)×102 |
| Листовое примордие | (2,9 ± 0,5)×102 | (1,5 ± 0,4)×102 | (8 ± 2)×10 | (4,1 ± 0,7)×102 |
| Семенные покровы | (5,0 ± 0,9)×102 | (9 ± 2)×10 | (1,0 ± 0,5)×10 | (7 ± 2)×10 |
Неоднородно распределение минеральных элементов и в эндосперме. В этом органе злаков различают ткань, обогащенную крахмалом, и алейроновый слой. Обычно алейроновый слой однорядный, клетки выглядят прямоугольными, однако у Hordeum sativum он трех- или четырехрядный. В целом форма, размеры, строение этого слоя варьируют у представителей различных подсемейств и триб злаков. В алейроновом слое сосредоточены макро- и микроэлементы, но содержание микроэлементов, как правило, на один-два порядка ниже, чем макроэлементов.
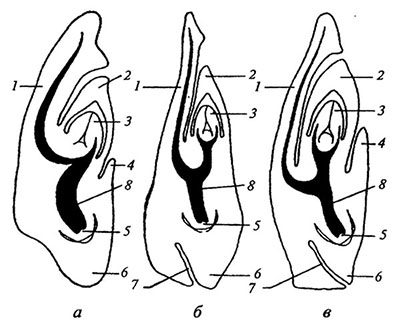
Рис. 4.4. Основные типы строения зародышевой злаков [модифицировано по: Жизнь растений, 1982].
а — фестукоидный (у ежи - Dactylis); б — паникоидный (у ветвянки — Brachiaria); в — эрагростоидный (у травы бизонов — Buchloe).
1 — щиток, 2 — колеоптиль, 3 — почечка, 4 — эпибласт, 5 — зародышевый корень, 6 — колеориза, 7 — щель между нижней частью щитка и колеоризой, 8 — проводящий пучок.
4.2.3. Формы
В семенах микроэлементы находятся в ионной и связанной форме. Металлы-микроэлементы в основном входят в состав фитатов — нерастворимых комплексов с различными катионами (Mg2+, Са2+, К+) фитиновой кислоты или миоинозитолгексафосфата

В семенах фитаты являются основным резервом фосфора. В зависимости от вида растений и органа семян содержание фосфора в фитиновой кислоте может варьировать от 40 до 86% общего фосфора. Фитин встречается главным образом в виде глобоидных включений внутри белковых тел, имеющих вакуолярное происхождение. При этом доля фитина в составе глобоидных кристаллов наиболее высока. Так, в глобоидах изолированных семян хлопка мало белков, жиров, углеводов, в то время как содержание фосфора составляет 14, 2%, фитиновой кислоты 60, а минеральных элементов 10%. В глобоидах зерновок риса содержится около 67% фитина, а семян Ricinus communis 78%. Глобоидные кристаллы по-разному распределены в белковых телах, их форма также разнообразна (призмы, друзы).
Зародыш, алейроновый слой, эндосперм, семядоли, щиток идентифицированы как органы биосинтеза и локализации фитина. Исключение из злаков составляет кукуруза, у которой до 88% фитиновой кислоты сосредоточено в зародыше. Концентрация фитина в кукурузе колеблется от 3 до 6 мкмоль на 1 г сырой массы зерновки. В формирующихся зерновках риса фитиновая кислота синтезируется в алейроновом слое и щитке, а в крахмалистом эндосперме не образуется. Вероятно, биосинтез фитина начинается в цистернах эндоплазматического ретикулума, затем фитин «упаковывается» в транспортные везикулы, которые мигрируют к вакуолярной мембране и сливаются с ней. На заключительных этапах биосинтеза частицы фитина накапливаются в полости вакуоли и формируют глобоид. Биохимические стадии синтеза фитиновой кислоты до конца не выяснены. Предположительно, сначала из глкжозы-6-фосфата формируется миоинозитол-1-фосфат, к которому присоединяются еще пять фосфорных групп (донор АТФ) и образуется миоинозитолгексафосфорная кислота (фитиновая кислота). Переходных форм от моно- к гексафосфорилированным формам миоинозитола не выявлено.
Ионы минеральных элементов присоединяются к фитиновой кислоте в результате взаимодействия с отрицательно заряженными фосфатными группами. Из минеральных элементов глобоидные кристаллы в наибольших количествах содержат фосфор, калий и магний, отмечаются также кальций, железо, марганец и другие микроэлементы. Глобоидные кристаллы из алейронового слоя пшеницы, удаленного от зародыша, содержат больше кальция, а вблизи зародыша больше железа.
Хотя минеральные вещества концентрируются главным образом в глобоидных кристаллах, некоторое количество элементов непосредственно связывается белками белковых тел. Из последних выделены, например, Mn-содержащие белки. К металлопротеинам относятся также лектины, которые кроме марганца могут содержать кальций.
Сведения о формах молибдена в семенах немногочисленны. Возможно, молибден связывается группой запасных белков, имеющих к нему высокое сродство. Часть этих белков представляет собой молибдокофактор (Мосо). В семенах небобовых (пшеницы, ячменя, табака, огурца и других культур) Мосо связан с двумя различными по молекулярной массе белками. Один из них, молекулярной массой около 200 кД, довольно устойчив, а другой, молекулярной массой около 60 кД, лабилен. Более высокомолекулярный белок, обнаруженный в зародышах пшеницы, обладает ксантиндегидрогеназной активностью. Белок 60 кД локализован в эндосперме и алейроновом слое семян и, по-видимому, представляет собой форму запасания кофактора.
Значительно содержание Мосо в семенах бобовых (горох, фасоль, люпин, соя). В покоящихся семенах гороха Мосо обнаружен в трех основных формах: термостабильных белков, термолабильных белков и диализируемого низкомолекулярного кофактора. Во всех выделенных формах Мосо находится в виде безмолибденового предшественника. Возможно, у бобовых молибден включается в Мосо при прорастании семян. Об этом свидетельствует появление активности ксантиндегидрогеназы и нитратредуктазы в процессе прорастания семян гороха одновременно с исчезновением в них Мосо-белков. Таким образом, Мосо-белки в семенах выступают донорами активного Мосо в процессе прорастания и, кроме того, предшественниками Мо-ферментов.
4.3. Прорастающие семена
Мобилизация органических и минеральных веществ семян в онтогенезе высших растений имеет ключевое значение. При низком содержании в среде элементов питания (особенно микроэлементов) минеральные резервы семени играют важную роль в росте и развитии растительного организма.
Например, наиболее информативным показателем обеспеченности растений молибденом служит его содержание в семенах (а не в вегетативных органах). Высокая эффективность применения молибденовых удобрений обеспечивается при содержании молибдена в семенах сои не выше 2,5 мг/кг. Если в зерновках злаков концентрация молибдена ниже 0,2-0,4 мг/кг, то у растений проявляются симптомы дефицита. При концентрации молибдена в семенах сои больше 5 мг/кг отпадает необходимость применения для этой культуры молибденовых удобрений. Критический для сои уровень марганца, ниже которого обнаруживаются симптомы дефицита, составляет около 20 мг/кг. Адекватные концентрации в семенах кобальта способствуют предотвращению заболевания люпина хлорозом при возделывании этой культуры без подкормок кобальтом.
При возделывании культурных растений на бедных (по микроэлементам) почвах в семенах содержится мало этих элементов питания. Посев таких семян на дефицитных по микроэлементам почвах приводит к слабому росту и низкому урожаю растений. Напротив, использование при посеве на почвах с низким плодородием обогащенного микроэлементами семенного материала обеспечивает больший прирост и урожай сельскохозяйственных культур. Для нормального прорастания семян пшеницы необходимо, чтобы концентрация в семенах, например, цинка была не меньше 5-10 мг/кг. Кроме того, растения, выросшие из семян с большим запасом микроэлементов, характеризуются лучшей устойчивостью к патогенам.
Содержание микроэлементов в семенах влияет не только на рост растений, но и на их элементный состав. Так, из зерновок пшеницы с высоким содержанием цинка развиваются проростки, характеризующиеся низкой скоростью поглощения корнями фосфора и калия. Такое влияние объясняется снижением конкуренции между микроэлементом и другими элементами питания в плазматической мембране на стадии поглощения этих элементов корнем. Интенсификация поглощения элементов при дефиците цинка в семенах может быть обусловлена активацией адаптивных механизмов растений, например выделением фитосидерофоров у видов стратегии II (см. гл. 6).
Необходимо отметить, что минеральные элементы могут не только поступать в формирующийся проросток из семян, но и откладываться в них. При нормальной обеспеченности среды питательными веществами часть поглощенных проростком макроэлементов аккумулируется в органах семян (эндосперм, семядоли). Например, поступление кальция из среды в побег и эндосперм зерновки кукурузы зависит от дифференцировки сосудов ксилемы корня. По мере формирования проводящей системы корня проростка может усиливаться приток кальция в органы зерновки. В случае достаточной обеспеченности среды минеральными элементами при сформировавшейся проводящей системе калий, кальций и фосфор эндосперма, вероятно, не играют существенной роли в питании проростка кукурузы.
Подвижность и доступность растениям многих микроэлементов, в отличие от макроэлементов, сильно ограничена, особенно в почвах карбонатных и высокоокультуренных. Поэтому в условиях агроценозов резерв микроэлементов в семенах, по-видимому, важнее для питания проростка, чем запасы макроэлементов. Снабжение же растений макроэлементами, особенно азотом и калием, зависит главным образом от внешних источников.
Интенсивная мобилизация запасных веществ в семенах обычно начинается после прорастания семян (рис. 4.5). При этом под прорастанием понимается цепь событий от поглощения покоящимися семенами воды до проникновения части зародыша (обычно зародышевого корешка) за пределы окружающих его структур семени. Усвоение формирующимися осевыми органами зародыша минеральных элементов зависит от донорно-акцепторных отношений между органами семени и проростка, а также от обеспеченности среды минеральными элементами. Кинетика поглощения проростком запасов минеральных элементов, сосредоточенных в семенах, определяется физиологическими потребностями зародыша, а также интенсивностью превращения (мобилизации) запасенных форм питательных веществ в растворимые. Мобилизация резервов минеральных веществ в семенах особенно важна для обеспечения проростка металлами-микроэлементами. В отличие от щелочноземельных (Са, Mg) и особенно щелочных (К) металлов 3d-элементы, главным образом железо и медь, а также цинк, характеризуются повышенной способностью образовывать устойчивые комплексы с органическими соединениями. Поэтому свободные формы, т. е. катионы этих металлов содержатся в растениях в небольшом количестве или вовсе не встречаются.
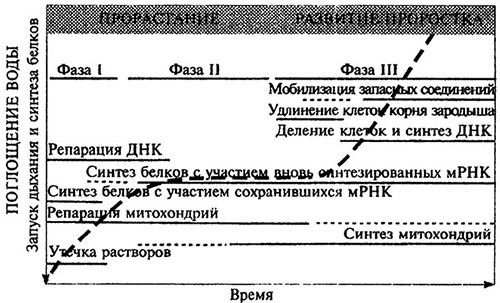
Рис. 4.5. Основные событие, происходящие при проростании семян и после их проростания [по: Bewley, 1997]
Динамика транспорта микроэлементов в прорастающих семенах наиболее детально изучена на примере зерновок кукурузы. Микроэлементы (Fe, Mn, Zn), как и кальций, в щитке, эндосперме и семенных покровах прорастающих зерновок кукурузы мобилизуются с разной интенсивностью (рис. 4.6). В течение первых пяти суток прорастания зерновок кукурузы на дистиллированной воде основной отток мобильных форм железа и марганца происходит из эндосперма, а цинка из щитка и (несколько меньше) из семенных покровов. Значительная мобилизация цинка из эндосперма регистрируется только после четырех и пяти суток инкубации зерновок на дистиллированной воде. Специфика мобилизации цинка объясняется, вероятно, особенностями распределения этого металла в зрелой зерновке. Высокая концентрация цинка отмечается в щитке и семенных покровах. При этом функциональные группы клеточных структур, участвующих в его связывании, повидимому, насыщены, что приводит к накоплению в щитке и семенных оболочках свободных и обменных форм цинка.
По мере роста проростка вклад эндосперма в формирование в зерновке общего фонда мобильных форм железа и марганца возрастает. Аналогичная зависимость отмечена и в отношении накопления в зерновке мобильных форм цинка за счет резервов щитка. Характерная особенность эндосперма растений различных видов состоит в том, что в ходе прорастания из этого органа довольно легко извлекаются многие минеральные вещества, как макро- (К, Mg, Са, Р), так и микроэлементы (Fe, Mn, Zn). Этот факт объясняют высокой проницаемостью тканей эндосперма.

Рис. 4.6 Динамика оттока микроэлементов и кальция из разных частей проростающих зерновки кукурузы
Как упоминалось выше (см. 4.2.3), в семенах значительное количество микроэлементов запасается в фитине, мобилизация которого зависит от активности фитаз и от реакции среды в фитатсодержащих вакуолях. Ацидофикация вакуолей растительных клеток важна при прорастании семян. Кислая реакция среды создает необходимые условия для деградации белка под действием кислых гидролаз, для гидролиза других соединений. Оптимум для работы фитаз также находится в кислой области pH. В результате исследований с использованием метода 31Р ЯМР-спектроскопии было установлено, что набухание зерновки кукурузы приводит к повышению мобильности фитина в вакуолях щитка зародыша. Растворимость фитата максимальна при минимальных значениях pH среды в зародыше, т. е. после трех-четырех суток прорастания зерновок. Вместе с тем фитазная активность в этот период крайне низка. Возможно, ацидофикация фитатов повышает мобильность этих соединений и усиливает их транспорт из фондов семян в побег и корни проростка.
Соответственно основная деструкция фитатных комплексов происходит в сформировавшихся в ходе прорастания вегетативных органах растения, а не в органах семян. Эта гипотеза основана на фактах аккумуляции фитаз в побегах и корнях пяти-восьмисуточных проростков кукурузы. Поэтому недостаточно высокий уровень ацидофикации в клетках зародыша и низкая активность фитазы, по-видимому, могут ограничивать мобилизацию в щитке нерастворимых форм микроэлементов, по крайней мере, в самом начале прорастания зерновок кукурузы.
Из расчетов следует, что даже в условиях голодания фонд образуемых в прорастающей зерновке кукурузы растворимых форм микроэлементов, а также кальция лишь частично используется растущими осевыми органами зародыша. Только через пять суток прорастания зерновки эффективность утилизации проростком растворимых форм металлов (Са, Fe, Zn) превышает 90%. Следовательно, на ранних этапах прорастания общий фонд микроэлементов и кальция, мобилизуемых во всех структурах зерновки, оказывается потенциально достаточным, чтобы обеспечить необходимое минеральное питание проростка. Вместе с тем существуют лимитирующие факторы: а) скорость мобилизации минеральных элементов в органах, выполняющих на том или ином этапе прорастания зерновки функции их основного источника; б) пространственная разобщенность зародышевой оси (акцептор) и запасов минеральных веществ в зерновке (донор). У злаков наибольшее влияние этих факторов, вероятно, приходится на начальные этапы прорастания зерновки, когда физиологическая активность ее структур (щитка, алейрона), участвующих в трансформации и транспорте запасных веществ, относительно невысока.
На самых ранних этапах прорастания (до двух суток) щиток по своим потенциальным возможностям может выполнять функцию основного донора растворимых форм некоторых микроэлементов (Fe, особенно Zn) для зародышевой оси. Вместе с тем в первые сутки прорастания образование мобильных форм отдельных микроэлементов в щитке мало связано, как обсуждалось выше, с трансформацией в нем нерастворимых соединений этих металлов, обусловленной работой фитаз и/или ацидофикацией фитатсодержащих компартментов. Основной отток этих микроэлементов происходит благодаря легкообменным формам, накопленным в период созревания зерновки на материнском растении. В связи с этим резервирование в зародыше мобильных форм микроэлементов в период созревания зерновки может рассматриваться как путь формирования генеративных органов у злаков, направленный на оперативную оптимизацию минерального питания растений на их ранних этапах развития. Создание в зародыше фонда мобильных форм микроэлементов — сильных комплексообразователей (особенно металлов), по-видимому, достигается за счет их интенсивного концентрирования в тканях. Высокие концентрации микроэлементов способствуют насыщению функциональных групп соединений растительной клетки, образующих комплексы с микроэлементами, и появлению их относительно подвижных форм.
Уровень концентрации микроэлементов в зародыше важен для роста проростка на гетеротрофной стадии развития. Например, концентрация в зародышах кукурузы различных генотипов железа и цинка коррелирует со скоростью роста корней.
Эффективность использования осевыми органами проростка мобилизованных в прорастающей зерновке микроэлементов зависит от локализации этих веществ. На ранних этапах прорастания эффективнее используются микроэлементы, локализованные в зародыше. У злаков наибольшее количество минеральных элементов зародыша сосредоточено в его самой массивной части — щитке. Сухая масса щитка кукурузы достигает 90% всей массы зародыша. Щиток является специализированным органом зерновки злаков, который запасает питательные вещества, выполняет транспортную и ацидофицирующую функции, осуществляет связь между эндоспермом и зародышевой осью. По существу, щиток представляет собой единственную семядолю зерновок злаков, которая образуется в результате асимметричного деления клеток предзародыша. По этой причине вторая семядоля остается в зачаточном состоянии. В зрелой зерновке щиток дорсальной стороной прилегает к эндосперму, а вентральной охватывает зародыш.
Основную массу щитка составляют паренхимные клетки, имеющие многоугольноокруглую форму. Цитоплазма клеток паренхимы содержит многочисленные липидные капли и белковые тела. Запасные вещества паренхимных клеток исчезают в первые три дня после начала прорастания зерновок. С осевыми органами щиток связан единой проводящей системой (рис. 4.4), что благоприятствует транслокации из него элементов питания. Позднее, по мере возрастания потребностей осевых органов в минеральных элементах и истощения запасов их растворимых форм в щитке, основное минеральное питание проростка осуществляется вследствие мобилизации резервов эндосперма, в которой главная роль принадлежит щитку и алейроновому слою.
В конце XIX в. было известно, что щиток, выполняя секреторную и всасывающую функции, участвует во внеклеточном переваривании крахмалистого эндосперма злаков. Мобилизация запасных веществ эндосперма осуществляется благодаря ацидофицирующей способности щитка. По данным С. М. Щипарева с коллегами (1976), секреция протонов щитком начинается на ранних стациях набухания зерновок. В результате после первых 14 ч прорастания зерновок кукурузы pH гомогенатов эндосперма снижается с 5,6 до 4,8. Подкисление эндосперма происходит неравномерно. Волна подкисления начинается от зоны непосредственного контакта эндосперма со щитком. В этой же зоне происходит наиболее интенсивное растворение эндосперма. В средах с изолированными щитками увеличивается концентрация многих органических кислот: глюконовой, лимонной, уксусной, пропионовой, масляной. Функциональная роль этих кислот в зерновке при прорастании не совсем ясна, однако их кислотные, а также хелатирующие свойства могут способствовать растворению резервных форм микро- и макроэлементов в эндосперме.
Секреция Н+ щитками — активный процесс, требующий затрат энергии. Подавляют секрецию протонов анаэробиоз и дыхательные яды (малонат, фторид натрия, динитрофенол), стимулируют катионы К+ и Са2+. Конкретные механизмы ацидофицирующей деятельности щитка изучены к настоящему времени недостаточно. Вероятно, в этом процессе участвуют Н+-АТФазы и окислительно-восстановительные цепи в плазмалемме эпителиальных клеток щитка. Ингибиторы транспортных АТФаз плазмалеммы (ортованадат, ДЭС, ДЦКД) подавляют выделение протонов щитками кукурузы на 20-40%. С помощью гистохимического метода установлено, что изолированные щитки кукурузы восстанавливают феррицианид, который служит индикатором редокс-цепей, причем наибольшая феррицианидредуктазная активность отмечается на поверхности щитка, обращенной к эндосперму. Восстановление феррицианида сопровождается усилением подкисления щитками внешней среды, что свидетельствует о выполнении редоксцепью функций Н+-помпы. Выделение кислот щитками стимулируют фитогормоны: ИУК и особенно кинетин в сочетании с ИУК. Фитогормоны также положительно действуют на секрецию щитками белков и поглощение сахаров и аминокислот.
Подкисление эндосперма может происходить и за счет углекислого газа, выделяемого щитками. Для щитков характерна высокая интенсивность дыхания, а их дыхательный коэффициент больше единицы (1,5-1,8). Наибольшая активность декарбоксилирующих ферментов отмечена в дорсальной части щитка, прилегающей к эндосперму.
Ацидофикация эндосперма опережает активную секрецию в него а-амилазы и других гидролаз. Функциональное значение такого опережения состоит в оптимизации реакции среды для работы кислых гидролаз. Кроме того, подавляется активность белковых ингибиторов амилазы и усиливается работа систем эпителиальных клеток, ответственных за транспорт продуктов гидролиза эндосперма.
На ранних этапах прорастания зерновок пониженная всасывающая способность щитка может лимитировать поглощение им минеральных веществ из эндосперма. По мере роста растений всасывающая функция щитка и соответственно поглощение минеральных веществ из эндосперма, вероятно, усиливаются.
В алейроновом слое эндосперма резервы (в том числе минеральные) и ферменты для их утилизации сосредоточены в вакуолях. Клетки алейронового слоя секретируют различные гидролитические ферменты. Это р-глюконаза и ксилоназа, которые разлагают клеточные стенки эндосперма, а-амилазы, другие гликозидазы, протеазы, пептидазы, кислые фосфатазы, ДНКазы, РНКазы, последовательно разрушающие содержимое клеток эндосперма. Деградация эндосперма начинается вблизи зародыша, а затем распространяется по всей зерновке, что сопровождается увеличением активности а-амилазы в призародышевой части эндосперма.
Поскольку для работы гидролаз оптимален узкий (кислый) интервал pH, активность этих ферментов регулируется в основном реакцией среды. Низкий уровень pH не только активирует работу вакуолярных гидролаз, но и создает электрохимический Н - градиент в тонопласте, способствующий вторичному транспорту резервов вакуоли в цитозоль. При этом аминокислоты, образующиеся вследствие гидролиза запасных белков вакуолей алейрона, используются для синтеза гидролаз de novo. Неорганические ионы, появляющиеся при разрушении вакуолярного фитина алейрона, поступают в эндосперм.
У растений известны два типа Н+-переносящих фосфогидролаз в тонопласте: вакуолярная АТФаза (В-АТФаза) и вакуолярная пирофосфатаза (В-ПФаза). В то время как В-АТФаза обнаружена в эндомембранах всех эукариот, В-ПФаза имеется только у растений.
Активность этих ферментов регулируется фитогормонами. Гиббереллины (GA3) стимулируют вакуолярную ацидофикацию алейрона, тогда как абсцизовая кислота (АБК) не оказывает значительного влияния на этот процесс. После шестичасовой инкубации в растворах GA3 значения pH среды в вакуолях клеток алейрона ячменя могут снижаться от 6,7-7,0 до 5,6-5,7. Механизм стимулирующего действия гибберелинов на ацидофикацию алейрона связывают скорее с активацией работы Н+-помпы, чем с увеличением количества белков помпы в вакуолярной мембране. В-ПФаза не задействована в стимуляции подкисления среды вакуолей при обработке GA3. Гибберелины повышают концентрацию цитоплазматического Са2+ в клетках алейрона злаков с 50-200 до 300-600 нмоль/л. В то же время активность В-ПФазы ингибируется Са2+, но ингибирующий эффект Са2+ устраняется высокими концентрациями Mg2+, запасы которого содержатся в вакуолях этих клеток. Поэтому в клетках алейрона регуляция В-ПФазы катионами может быть комплексной. Вопросы влияния гибберелинов на изменение концентрации кальция в вакуоли требуют дальнейшего изучения.
Гидролитическую активность алейронового слоя жестко контролирует рост зародыша. Посредниками в этом процессе служат фитогормоны, в основном гибберелины и абсцизовая кислота, известные как антагонисты. Гиббереллины, секретируемые зародышем, усиливают, а АБК подавляет синтез и активность гидролаз алейроновым слоем. Регуляция фитогормонами гидролитической активности алейрона тесно связана с регуляцией концентрации ионов кальция и pH в цитозоле, а также с экспрессией гена α-амилазы алейронового слоя. Индуцируемые гибберелинами высокая концентрация Са2+ в цитозоле и низкое значение pH благоприятны для секреции и активации гидролаз. АБК, напротив, вызывает понижение концентрации ионов кальция в цитозоле и повышение значения pH в клетке.
После завершения секреторных функций клетки алейрона отмирают. Их гибель рассматривается как запрограммированная (апоптоз). Характерные черты апоптоза — сжатие ядра, конденсация хроматина, разрыв ДНК. Процесс апоптоза эволюционно консервативен и играет существенную роль в эмбриогенезе и развитии. У растений апоптоз наблюдается под действием стресса и грибной инфекции. При прорастании зерновки наиболее интенсивная фрагментация ДНК происходит в частях алейрона вблизи зародыша. В запуске апоптоза могут участвовать гибберелины, выделяемые зародышем. Апоптоз клеток алейрона, вероятнее всего, обусловлен, по аналогии с клетками млекопитающих, интенсивной секрецией в нем гидролаз, особенно протеаз, под влиянием гибберелинов зародыша. Осмотический стресс, возникающий в зерновке при накоплении в ней продуктов деградации крахмала, не рассматривается в качестве возможного триггера апоптоза. Возможно, при неблагоприятных условиях вступление алейрона в состояние апоптоза предотвращает АБК через регулирование активности и синтеза гидролаз, например в результате изменения концентрации кальция и/или pH в клетке.
Эффективность перераспределения минеральных элементов между органами прорастающего семени и проростка варьирует в зависимости от вида растений и природы элемента. Из семядолей в осевые органы развивающегося люпина может поступать до 90% фосфора, калия и серы, до 60-90% магния, железа, цинка, марганца, но только 25-30% кальция. Пастбищные травы и бобовые используют семенные резервы фосфора, железа и серы более эффективно, чем азота и калия.