

В главе дана общая характеристика процессов комплексообразования и химических свойств комплексов. Приведены примеры наиболее важных комплексов микроэлементов в растениях и почвах. Рассмотрена специфика усвоения комплексонатов микроэлементов корнями и побегом растения. Идентифицированы свойства комплексов, определяющие эффективность их усвоения растительным организмом. Описаны основные принципы подбора физиологически активных форм комплексонатов микроэлементов при проведении корневых и некорневых подкормок культурных растений.
5.1. Свойства комплексов
5.1.1. Общие сведения. Химические свойства
Большей части микроэлементов в силу их физико-химических свойств присуща очень высокая реакционная способность к образованию комплексов (комплексонатов) с органическими веществами. Не случайно поэтому в растениях основное количество микроэлементов представлено в виде комплексных соединений. В почве комплексообразование также играет важную роль, определяя в значительной степени мобильность и доступность соединений микроэлементов растениям. Рассмотрение свойств комплексов необходимо для понимания многих процессов, происходящих с участием микроэлементов в системе растение-почва.
Первоначально термином комплекcоны, предложенным немецким ученым Г. Шварценбахом (1945), называли органические лиганды группы полиаминполиуксусных кислот, в составе которых находятся иминодиацетатные фрагменты, связанные с алифатическими и ароматическими радикалами. Позже к комплексонам стали относить аналогичные по строению и свойствам соединения с более широким спектром функциональных групп: алкилфосфоновые, алкиларсоновые, алкилсульфоновые, и атомов: фосфор (III), сера (II), селен (II), теллур (II). В настоящее время термином комппексоны обозначают обширную группу органических веществ с кислыми и основными донорными центрами, образующих с катионами металлов не менее двух замкнутых циклов. В зарубежной литературе большее распространение получили термины «хелаторы», «хелатирующие агенти». Термином хелат (от греческого chele — клешня), предложенным Г. Морганом (1920), или комплексонатобозначают циклические структуры, которые образуются в результате присоединения катиона к двум или более донорным атомам, принадлежащим одной молекуле хелатирующего агента.
Способность при взаимодействии с микроэлементами образовывать устойчивые циклические структуры физиологи растений рассматривают как наиболее важное свойство комплексонов. В реакциях комплексообразования участвуют такие структурные компоненты комплексонов как донорные атомы и их попарные связки — «клешни». Образование хелатного комплекса, например с микроэлементом, происходит только тогда, когда катион одновременно касается того и другого донорного атома. При этом хорда, соединяющая два соседних атома «клешни», не должна пересекать никаких других химических связей, а ее длина не должна превышать 0,4 нм. К центральному атому лиганды присоединяются двуцентровыми σ-, π-, δ-связями и многоцентровыми связями. Химическая связь между центральным ионом и лигандом образуется только при условии соответствия симметрии орбиталей металла (М) и лиганда (L), например, донорные орбитали (а или я) взаимодействуют с аналогичными акцепторными. При этом центральный ион может выступать в роли как акцептора, так и донора электронов.
Число атомов или их групп в молекуле лиганда, координируемое центральной частицей в данном соединении, называется координационным числом (КЧ). Если связи между ядром и лигандом двуцентровые, то КЧ равно числу образуемых центральной частицей связей, т. е. числу непосредственно связанных с ней донорных атомов лиганда. В обобщенном виде реакция образования хелата с участием металла (M) и лиганда (L) может быть представлена в следующем виде:
[M(H2O)x]n++Lm- ↔ [M(H2OyL]n-m+(x-y) H2O
Количественной характеристикой этого равновесия служит термодинамическая константа устойчивости (β°)
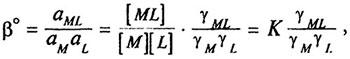
где α — активность иона, γ — коэффициент активности иона. Термодинамическая константа равновесия — мера изменения стандартной свободной энергии Гиббса ΔG°, энтальпии ΔН° и энтропии ΔS° комплексообразования:
-RT In β° = ∆G°=∆H° - T∆S°
Определение коэффициентов активности ионов технически затруднено. Поэтому для количественной оценки реакции комплексообразования вместо термодинамических констант используют стехиометрические (концентрационные) константы устойчивости (К), выражающие отношение не активностей, а концентраций реагентов:
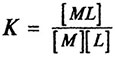
Сравнение концентрационных констант устойчивости между собой допустимо при одинаковой величине ионной силы раствора (р), создаваемой одним и тем же электролитом.
При расчете условных констант устойчивости вводят поправку на побочные реакции, например образование протонированных форм лиганда (НL, Н2L ... НхL), гидроксокомплексов (М(ОН)L), гидролиз ионов металла (M(ОН)х), протонированных комплексов (MHL):
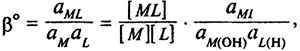
где аL(H), aM(OH), aML— коэффициенты активности побочных реакций протонирования лиганда, образования гидроксокомплексов и гидроксокомплексонатов. Условная константа устойчивости является функцией pH и позволяет определять оптимальный интервал pH, при котором происходит комплексообразование с определенным катионом.
Важнейшее свойство хелатов — устойчивость к диссоциации на металл и лиганд, зависит от многих сопряженных между собой факторов, идентификация которых в чистом виде весьма затруднена. Наибольшее значение имеют природа хелатора и катиона. Существенное влияние на прочность хелатов оказывают гибкость молекулы лиганда, величина ее заряда, состав и число донорных центров, а также способы их сочленения в молекуле лиганда. К наиболее существенным характеристикам центрального иона относят степень его окисления и размер, строение электронных оболочек. Широкий диапазон изменения значений перечисленных параметров затрудняет выявление общих закономерностей устойчивости комплексных соединений, весьма разнообразных по составу. Вместе с тем для отдельных групп комплексонатов все же установлен ряд параметров, определяющих устойчивость хелатных комплексов: дентатностъ лиганда, размер хелатного цикла, электростатический фактор.
Дентатностъ лиганда. Под дентатностью, или координационной емкостью подразумевают число донорных атомов комплексона, образующих в данном соединении координационные связи. Иными словами, речь идет о количестве мест, которые занимает лиганд в координационной сфере конкретного хелатного комплекса. Истинная дентатностъ, проявляемая комплексоном в водном растворе по отношению к катиону, известна для ограниченного числа соединений. Обычно выражают зависимость Ig КML от потенциальной дентатности лиганда, рассматривая каждую донорную группу или атом как монодентатные. При сравнении констант устойчивости в ряду однотипных лигандов установлено, что с увеличением дентатности комплексонов устойчивость их хелатов, как правило, повышается до насыщения координационной емкости.
Размер хелатного цикла. Реакция комплексообразования между металлом и лигандом сопровождается замыканием циклов, одним из звеньев которых выступает ион металла (М). Устойчивость индивидуального хелатного цикла зависит от его размера и напряженности. Наибольшей устойчивостью характеризуются пяти- и шестичленные циклы.
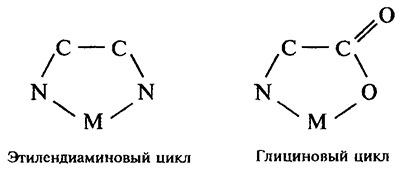
Четырехчленные хелатные циклы наименее устойчивы. Низкая устойчивость последних обусловлена в основном стерическими эффектами, в частности увеличенной напряженностью валентных углов, измененной длиной связи и конформацией. Циклы с числом звеньев выше шести, как правило, малоустойчивы.
Электростатический фактор. В реакциях комплексообразования практически всегда действуют силы, которые рассматривают как чисто ионные. С возрастанием напряженности поля центрального катиона, увеличением его заряда и уменьшением радиуса устойчивость хелатных комплексов повышается. Донорные атомы с большими значениями отрицательного заряда характеризуются большим сродством к катионам. Поэтому протонированные комплексы, как правило, менее устойчивы к диссоциации по сравнению с их нормальными формами. Следует, однако, отметить, что соответствие параметров (донорно-акцепторных, стереохимических и др.) центрального иона и лиганда приводит к эффективному конкурированию ионов металлов с протонами за донорные атомы, которые в отсутствие ионов металлов в водных растворах не депротонируются (атом азота в пептидах) или депротонируются в сильно щелочной среде (пиррольный азот в имидазоле, гистидине, гетероатомы азота в пуриновых и пиримидиновых основаниях нуклеотидов). В результате рК депротонирования может снижаться на несколько порядков. Конкуренция металлов (Pb2+ > Pt2+ > Cu2+ > Ni2+ > Со2+ > Zn2+) с Н+ за азот пептидной группы снижает pH депротонизации с 13 до 3-6. Образование комплексов триптофана с Сu2+ может происходить даже в кислой среде (pH < 3), хотя в отсутствие этого иона его рК превышает 13. Понижение эффективного заряда катиона происходит и при появлении ионов гидроксила в составе комплексоната, что способствует его распаду.
Катионы переходных 3d-металлов, к которым относится значительное число микроэлементов, образуют в двухвалентном состоянии нормальные комплексы со многими комплексонами. Для большинства лигандов при обычных условиях сохраняется единый порядок изменения устойчивости комплексонатов (ряд Ирвинга-Вильямса):
Са2+ < Mg2+ < Mn2+ < Fe2+ < Со2+ < Zn2+ < Ni2+ < Cu2+.
Изменения констант устойчивости комплексов микроэлементов симбатны изменению энергии их 3d-атомных орбиталей.
В отличие от многих микроэлементов щелочноземельные и особенно щелочные элементы обладают значительно меньшей комплексообразующей способностью. Не случайно поэтому калий в растениях в основном содержится в виде свободных ионных форм. Изменение степени окисления 3d-элементов сопряжено с существенным уменьшением или увеличением устойчивости хелатов. Например, трехвалентные катионы марганца, железа, кобальта образуют с одними и теми же лигандами комплексы приблизительно на 10 порядков более устойчивые, чем двухвалентные катионы этих металлов.
Изменение состава донорных атомов и строения лигандного контура существенным образом влияет на общую устойчивость комплексонатов и на относительную селективность действия хелаторов на микроэлементы. Под селективностью подразумевается способность молекулы лиганда формировать комплексы с определенными ионами центральных атомов. Обычно аминокарбоновые комплексоны образуют более устойчивые комплексы, чем, например, лиганды с фосфоновыми группами. Однако селективность последних в раде случаев выше, чем селективность первых. Введение в контур лиганда серы вместо азота резко снижает значения KML и нивелирует различия между катионами переходных 3d-элементов.
Электростатический фактор не относится к определяющим при образовании комплексов с ковалентной связью, однако и в этом случае с увеличением степени окисления катионов устойчивость их комплексов, как правило, возрастает.
5.1.2. Комплексы микроэлементов в почвах и растениях
В почвах, особенно в их верхних корнеобитаемых горизонтах значительное количество микроэлементов находится в форме комплексных соединений с гуминовыми веществами. Химическая природа гуминовых веществ окончательно не установлена, что обусловлено их чрезвычайно сложной молекулярной структурой. По мнению большинства исследователей, гуминовые вещества представляют собой полимеры с широким интервалом молекулярно-массового распределения, их структурные фрагменты состоят из ядра ароматической природы и нерегулярных периферических открытых цепей. Длина соединительных цепей между структурными единицами в большинстве случаев соответствует сумме трех-пяти углерод-углеродных связей. В реакциях комплексообразования с металлами принимают участие карбоксильные и карбонильные функциональные группы, фенольные и спиртовые гидроксилы, азотсодержащие и другие структурные фрагменты гуминовых веществ. Высокая обменная емкость гумусовых веществ главным образом обусловлена карбоксильными и фенольными оксигруппами в их составе, выполняющими основную роль в образовании комплексов с ионами металлов.
Сведения об устойчивости соединений металлов с органическим веществом почвы весьма противоречивы. Приведем ориентировочные интервалы значений констант устойчивости (IgK) для гуминовых веществ [по: Орлов, 1990], где минимальные величины IgK измерены при pH 2-4, максимальные при pH 8-10.
| Гуматы | lg К | Фулъваты | lg К | Фулъваты | lg К |
| ГК-Fe | 2,4-9,8 | ФК-Mg | 1,2-2,1 | ФК-Со | 2,2-3,7 |
| 7,0-9,7 | ФК-Аl | 6,5 | ФК-Ni | 3,5-4,1 | |
| ГК-Zn | 2,9-10,8 | ФК-Са | 2,0-2,9 | ФК-Сu | 3,2-8,7 |
| ГК-Cd | 6,9 | ФК-Мn | 1,5-3,8 | ФК-Zn | 1,7-9,3 |
| ГК-Pb | 8,7 | ФК-Fe | 5,1-6,1 | ФК-Рb | 3,1-6,1 |
Величины констант устойчивости варьируют в зависимости от почвенных условий, метода измерения и выбранных значений молекулярных масс. С повышением pH устойчивость органоминеральных комплексов, как правило, возрастает.
Определение строения соединений, образуемых в результате взаимодействия гуминовых веществ с металлами, входит в число наиболее трудных задач почвоведения. Органоминеральные комплексы почв весьма разнообразны по форме. Идентифицированы как внутри-, так и внешнесферные комплексы с металлами. В результате даже простые ряды сродства катионов к гуминовым кислотам могут существенно варьировать в зависимости от свойств почвы и методов исследования. Поэтому для прогнозирования взаимодействия металлов с гуминовыми веществами рекомендуется пользоваться частными рядами устойчивости органоминеральных комплексов, установленными для конкретных почв. Приведем примеры возможных взаимодействий металлов с гуминовыми и фульвокислотами (рис. 5.1).
Основной миграционной формой многих микроэлементов в почвах и природных водах являются относительно прочные растворимые фульватные комплексы анионного типа. Эти соединения играют важную роль в минеральном питании растений. В результате экспериментов с 59Fe, например, было установлено, что при оптимальных концентрациях доступность растениям Fe-фульватных комплексов сопоставима с доступностью Fe-цитрата.
Во взаимодействии с металлом участвуют амино- и карбоксильные группы, а также группы, встречающиеся в боковых цепях, например в серине и тирозине ОН-, в цистеине SH-группа. Некоторые аминокислоты, имеющие в боковой цепи электрондонорную группу, способны играть роль тридентатных лигандов по отношению к металлу. Форма и размеры боковой цепи определяют пространственные эффекты в реакциях хелатирования.
Пептиды представляют собой производные аминокислот, образующиеся в результате конденсации двух и более аминокислот, сопровождающейся выделением воды. В пептидах донорами электронной пары для иона металла могут быть кроме аминной и карбоксильной групп азот пептидной связи и сера. Реакции комплексообразования с участием пептидов играют важную роль в механизмах устойчивости растений к действию высоких концентраций металлов.

Рис. 5.1. Вероятные механизмы взамодействия гуминовых и фульвокислот с металлами [Schnitzer, 1986, цит. по: Орлов, 1990]
Известны две основные группы пептидов, способных существенно снижать фитотоксичность металлов: металлотионеины и фитохелатины.
Металлотионеины представляют собой низкомолекулярные полипептиды, богатые цистеином, он и обладает высоким сродством к меди. Этот металл индуцирует образование металлотионеинов. Уровень экспрессии гена МТ 2, кодирующего синтез металлотионеинов, коррелирует с устойчивостью растений к меди.
Фитохелатины также относятся к группе низкомолекулярных пептидов, обогащенных цистеином. Они могут участвовать в детоксикации не только меди, но и кадмия.
В белках донорные атомы для ионов металлов такие же, как и в пептидах. Строение важнейших ферментных белковых комплексов рассмотрено выше (см. 3.2).
В последнее время интенсивно изучаются реакции комплексообразования с участием нуклеиновых кислот и нуклеотидов. В формировании хелатных структур могут принимать участие атомы азота азотистых оснований, кислород фосфатных групп и гидроксильных групп рибозы. При взаимодействии катионов металлов с фосфатными группами возможно формирование структур нескольких типов:
- Хелатный комплекс с двумя атомами кислорода одной фосфоновой группы, формирование которого сопровождается образованием четырехчленных циклов. Такие циклы довольно устойчивы только с катионами большого радиуса:

- Восьмичленные циклы, образующиеся в случае взаимодействия катиона металла с атомами кислорода двух фосфатных групп одновременно, которые могут упрочняться внутримолекулярными связями с участием молекул воды:

- Полимерные структуры, в которых лиганд служит мостиком между катионами. В этой реакции комплексообразования могут быть задействованы все три атома кислорода фосфатной группы.
Важная роль в регуляции поглощения и транспорта микроэлементов в системе растение—почва принадлежит комплексам микроэлементов с органическими кислотами. В реакциях комплексообразования участвуют карбоксильные, спиртовые и кетогруппы органических кислот. Информация о влиянии органических, а также аминокарбоновых кислот на питание растений микроэлементами приведена ниже (см. гл. 6).
5.2. Усвоение комплексов микроэлементов растениями
5.2.1. Корень
Комплексонаты (хелаты) рассматривают как одну из основных форм содержания микроэлементов в ризосфере корня (см. гл. 6). Поэтому целесообразно рассмотреть наиболее важные для минерального питания растений свойства хелатов. Приведем структурные формы некоторых синтетических хелаторов (рис. 5.2) и константы устойчивости их комплексов с металлами (табл. 5.1). Основные закономерности усвоения корнями растений комплексов микроэлементов установлены с использованием природных и синтетических Fe-хелатов, многие из них широко используются в растениеводстве при возделывании растений на карбонатных почвах и в условиях гидропоники.
Комплексы железа
Влияние комплексонов на корневое питание растений железом представляет собой сложный процесс, биологическая эффективность которого определяется множеством факторов, прежде всего обеспеченностью растений железом. При нормальном содержании железа в питательном растворе природа лиганда практически не влияет на физиологическую эффективность Fe-комплексов, определяемую по действию хелатов на содержание хлорофилла в листьях и биомассу растений. Об этом свидетельствуют идентичные характеристики формирования биомаіхы и концентрация хлорофилла в листьях растений, которые выращены на растворах, содержащих разные Fe-хелаты в достаточном количестве (рис. 5.3).
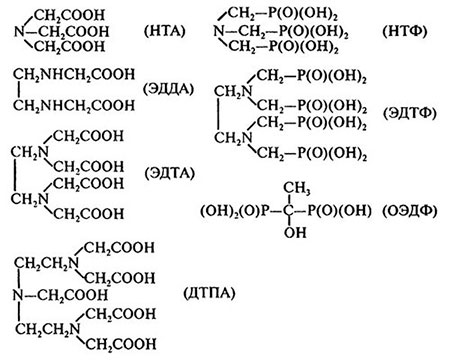
Рис. 5.2. Структурные формулы некоторых хелатирующих агентов
В условиях Fe-дефицита наблюдаются иные закономерности, которые имеют свою специфику в зависимости от реакции среды.
Кислая среда. В кислых средах наибольшая биологическая эффективность присуща фосфорсодержащим комплексам (Fe-ОЭДФ), а наименьшая аминокарбоновым (Fe-ДТПА и Fe-HTA) (рис. 5.4). При этом не выявлено корреляции между устойчивостью комплексов к диссоциации и их физиологической эффективностью. Разрушение Fe-хелатов в кислой питательной среде происходит с меньшей интенсивностью, в частности, вследствие снижения вероятности образования гидроксокомплексов железа и хелатов щелочноземельных (особенно Са) металлов. Поэтому достаточная для растений с различными типами (стратегия I, II) адаптации к Fe-дефициту (см. гл. 6) растворимость соединений железа может достигаться в кислых растворах даже при использовании Fe-хелатов с относительно невысокой устойчивостью. Вместе с тем при низкой концентрации в питательной среде хелатированного железа его усвоение корнем могут ограничивать силы электростатического отталкивания между одноименно заряженными Fe-комплексами и корневой поверхностью.
| Лиганд | Mg2+ | Са2+ | Мn2+ | Fe2+ | Fe3+ | Со2+ | Сu2+ | Zn2+ |
| НТА | 5,47 | 6,39 | 7,46 | 8,33* | 15,9 | 10,38 | 12,94 | 10,66 |
| ЭДДА | 3,95 | Нет данных | 7,05 | Нет данных | Нет данных | 11,25 | 16,2** | 11,22 |
| ЭДТА | 8,83 | 10,61* | 13,81 | 14,27 | 25,0 | 16,26 | 18,70 | 16,44 |
| ДТПА | 9,34 | 10,75 | 15,51 | 16,4 | 28,0* | 19,15 | 21,38 | 18,19 |
| НТФ | Нет данных | Нет данных | 10,2 | Нет данных | 14,60*/21,37 | 14,4 | 17,8 | Нет данных |
| ЭДТФ | 8,63 | 9,33 | Нет данных | » | Нет данных | 17,36 | 23,0 | 19,52 |
| ОЭДФ | 6,6 | 6,0 | 9,2 | 9,1 | 16,3 | 9,4 | 12,5 | 10,7 |
Примечание *Температура 20°С; **30°С
Следует отметить, что процессы физико-химической адсорбции играют важную роль в поглощении растением микроэлементов. Приоритет в аккумуляции катионов из почвенных растворов с низким содержанием ионов принадлежит клеточным оболочкам, участвующим в создании запасного фонда микроэлементов в клетке. Прочному удерживанию поверхностью корня катионов металлов способствует отрицательный заряд клеточных стенок, обусловленный депротонированными карбоксильными группами полисахаридов. Клеточные стенки способны накапливать катионы металлов. Наибольшее накопление на поверхности клеточных стенок катионов происходит при их относительно невысоких равновесных концентрациях в растворах (табл. 5.2). Незначительное содержание в клеточной оболочке карбоксильных групп создает предпосылки для высокой потребности культуры в микроэлементах. Накопление микроэлементов на поверхности корней может происходить в виде малодоступных для транслокации в надземные органы растений соединений, что весьма характерно для минеральных, солей железа, образующего нерастворимые в воде гидроксиды в широком диапазоне «физиологических» значений pH растворов.
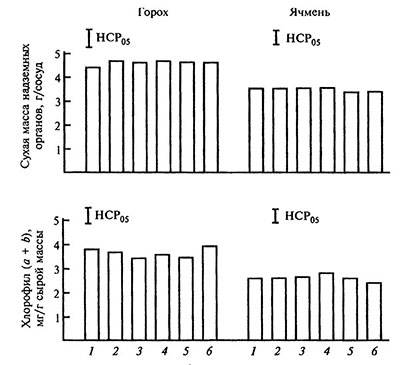
Рис. 5.3. Содержание хлорофила в листьях и формирование биомассы гороха и ячменя под влиянием хелатирующих агнетов разной природы при достаточной обеспеченности растений железом [по: Битюцкий, 1995].
Корневое питание, рН питательных растворов 5,6; Fe : L = 1:1; концентрация железа в опытах с ячменем 3,57×10-5, в опытах с горохом 7,14 ×10-5 моль/л.
1 — Fe-ДТПА, 2 — Fe-ЭДТА, 3 — Fe-НТА, 4 — Fe-ЭДТФ, 5 — Fe-НТФ, 6 — Fe-ОЭДФ
В составе хелатов катионные свойства металлов маскируются, что способствует снижению уровня их накопления на поверхности корня. Однако при малой концентрации металла в растворе электростатический фактор может лимитировать контакт корневых систем с обычно отрицательно заряженными молекулами хелата и нарушать процесс их метаболизации. На примере бора, ванадия, молибдена и других элементов установлено, что отрицательно заряженные поверхности клеточных стенок замедляют поглощение анионов микроэлементов из растворов с их пониженным содержанием. Возможно, различия в эффективности действия Fe-xeлатов в кислой среде обусловлены разными величинами их зарядов и, следовательно, их разной способностью адсорбироваться поверхностью корня.
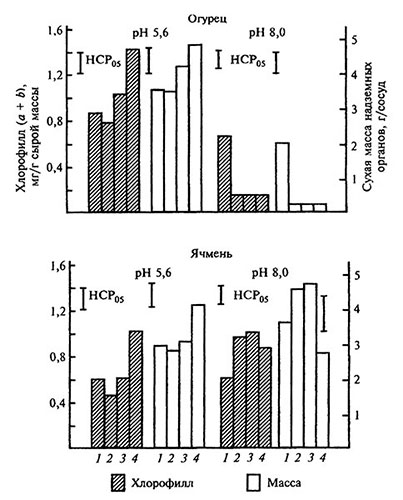
Рис. 5.4. Содержание хлорофилла в листьях и формирование биомассы растений огурца и ячменя при низкой обеспеченности железом под влиянием хелатириюущих агентов и рН среды [по: Битюцкий 1995].
Корневое питание, С=4,5×10-6 моль/л 1 — Fe-ДТПА, 2 — Fe-НТА, 3 — Fe-НТФ, 4 — Fe-ОЭДФ рН 8 создавали внесением NaHCO3 в раствор, рН 5,6
Как известно, заряд хелата определяется основностью органического лиганда, значениями его констант протонирования (рА), степенью окисления катиона. Карбоксильные группы характеризуются низкими значениями рК, что свидетельствует об их способности диссоциировать в сильно кислых растворах.
| Оболочки | Десорбция водой | 5Концентрация цинка, моль/л | |||
| 5•10-6 | 5•10-4 | ||||
| мг • экв /г | % | мг • экв /г | % | ||
| Горох | |||||
| Без пектиновых веществ | Раствор | 0 | 0 | 0,34 ± 0,06 | 6,4 |
| Остаток | 0,19 ± 0,02 | 100 | 4,93 ± 0,07 | 93,6 | |
| Без пектиновых веществ и гемицеллюлоз | Раствор | 0 | 0 | 0,09 ± 0,01 | 8,1 |
| Остаток | 0,08 ± 0,01 | 100 | 1,02 ± 0,11 | 91,9 | |
| Без карбоксильных групп | Раствор | 0 | 0 | 0,36 ± 0,02 | 2,5 |
| Остаток | 0,15 ± 0,04 | 100 | 1,09 ± 0,07 | 97,5 | |
| Кукуруза | |||||
| Без пектиновых веществ | Раствор | 0 | 0 | 0,48 ± 0,03 | 16,6 |
| Остаток | 0,24 ± 0,01 | 100 | 2,40 ± 0,07 | 83,4 | |
| Без пектиновых веществ и гемицеллюлоз | Раствор | 0,01 ± 0 | 8,3 | 0,20 ± 0,01 | 20,9 |
| Остаток | 0,11 ±0,01 | 91,7 | 0,76 ± 0,40 | 79,1 | |
| Без карбоксильных групп | Раствор | 0,01 ± 0 | 8,9 | 0,06 ± 0,01 | 14,8 |
| Остаток | 0,11 ±0,01 | 91,1 | 0,36 ± 0,02 | 85,2 | |
Поэтому при «физиологических» значениях pH с увеличением числа карбоксильных групп в молекуле хелатирующего агента будет возрастать не только устойчивость образуемых хелатов, но и их отрицательный заряд. В группе аминокарбоновых хелатов (рис. 5.2), широко используемых в практике растениеводства, максимальные величины отрицательного заряда отмечены для комплексов Fe-ДТПА и Fe-ЭДТА, содержащих в своем составе наибольшее число карбоксильных групп. Относительно высокий отрицательный заряд этих комплексов — основная причина их слабой абсорбции корнями (рис. 5.4), а следовательно, и слабого антихлорозного действия. На примере железофульватных комплексов также установлено, что наиболее доступны для растений (пшеница) положительно заряженные Fe-фульваты.
Два протона в составе фосфоновой группы обусловливают ее большую потенциальную основность. Так, нитрил триметиленфосфоновая (НТФ) и этилендиаминтетраметиленфосфоновая (ЭДТФ) кислоты — фосфорсодержащие аналоги трех- и четырехосновных нитрилтриуксусной (НТА) и этилендиаминтетрауксусной (ЭДТА) кислот, являются соответственно шести- и восьмиосновными кислотами. Однако фосфонoвые группы, в отличие от карбоксильных, характеризуются существенно более высокими значениями pH и повышенным сродством к протону. Например, гидроксиэтилидендифосфоновая кислота (ОЭДФ) четырехосновна, но диссоциации ее гидроксигруппы с отщеплением Н+ не наблюдается, по крайней мере до pH 13. Поэтому фосфоновые группы в составе лиганда, особенно если их немного, как например у ОЭДФ, создают благоприятные условия для формирования Fe-хелатов с низкими отрицательными зарядами, что способствует лучшей абсорбции корнем железа в функционально активной форме.
О влиянии лигандов на функциональную активность абсорбированного железа можно судить по результатам экспериментов с исключением из питательных растворов Fe-хелатов после их кратковременного взаимодействия с корнями растений (рис. 5.5). В период после исключения Fe-комплексов из растворов проявление признаков хлороза у растений происходит с аналогичной описанной выше закономерностью: поражение растений хлорозом больше в вариантах с Fe-ДТПА, Fe-ЭДТА, Fe-ЭДТФ, меньше в вариантах с Fe-ОЭДФ и Fe-НТФ. Следовательно, устойчивость к диссоциации и способность к адсорбции корневыми системами — это основные свойства Fe-комплексонатов, способствующие оптимизации усвоения растением железа при условии его ограниченного содержания в кислой среде.
Сочетание этих свойств трудно достижимо в ряду аминокарбоновых лигандов, так как депротонизация необходима для участия карбоксилов в образовании координационных связей с металлами. Поэтому увеличение числа карбоксильных групп в молекуле комплексона ведет к повышению устойчивости комплекса и его отрицательного заряда. Кроме того, наращивание звеньев в комплексе целесообразно до определенною предела, соответствующего насыщению координационной емкости. Дальнейшее увеличение размеров хелатного цикла может сопровождаться ослаблением прочности комплексоната, что весьма характерно для хелатов Fe (III). Содержание в среде фосфорсодержащих Fe-хелатов типа ОЭДФ с минимально допустимым для формирования хелатных структур количеством фосфоновых групп более благоприятно для минерального питания высших растений.
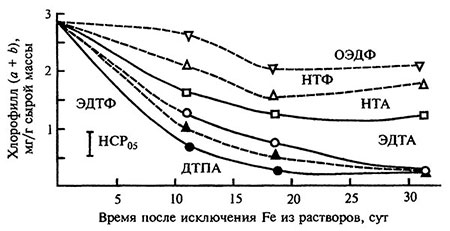
Рис. 5.5. Сожержание хлорофила в листьях ячменя после кратковременного действия Fe-хелатов.
Вораст растений 10-41 день. На питательном растворе с Fe-хелатами (Сi•c=3,57•10-5 моль/л, рН 5,6) растения выращивали с 5- до 10-го дня, на растворе без Fe-хелатов с 10- до 41-го дня.
Щелочная среда. В щелочной среде, особенно при повышенных концентрациях металлов, конкурирующих за места связывания в комплексе, преимущество принадлежит органическим лигандам с высокой селективностью к железу и способностью формировать высокоустойчивые хелаты. Это свойство хелатов особенно важно для растений, произрастающих на карбонатных почвах со щелочной реакцией среды и высоким содержанием кальция. Конкуренция кальция за лиганд может приводить к разрушению относительно неустойчивых Fe-комплексов и образованию плохо растворимых в воде гидроксидов железа (рис. 5.6). Физиологическое значение перечисленных свойств Fe-хелатов выше для растений стратегии I (см. гл. 6). Как известно, растения стратегии I адаптируются к Fe-дефициту с помощью выделения корнями Н+ и активации Fe-редуктаз.
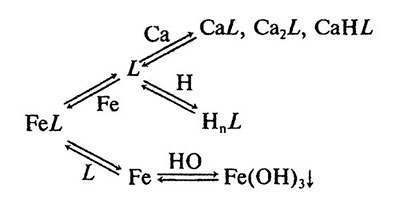
Рис. 5.6. Равновесные реакции в системе Са-Fe-L [по: Lucena et al., 1988]
Основной путь адаптации к недостатку железа растений стратегии II связан с продуцированием корнями фитосидерофоров. (Более подробная информация по этому вопросу содержится в гл. 6.) Для ячменя (стратегия II) рост растений и содержание хлорофилла в листьях были выше на растворах (щелочная среда) со среднеустойчивыми хелатами, а для огурца (стратегия I) с высокоустойчивыми (рис. 5.4). При этом для огурца обнаружена сильная прямая корреляция содержания хлорофилла в листьях, а также биомассы с концентрацией в растворах водорастворимых форм железа.
Эти видовые особенности усвоения корнями Fe-хелатов в щелочных средах объясняются спецификой механизмов адаптации различных групп растений к Fe-дефициту. У растений стратегии I функционирование Fe-редуктаз и продуцирование протонов оптимально при pH 5 и подавляется в щелочной среде, а также в случае, если в корневой зоне нет подвижных форм железа. Фитосидерофорный механизм адаптации (стратегия II) в меньшей степени подвержен негативному воздействию высоких значений pH среды, что позволяет злакам эффективно усваивать железо, даже если в растворе практически нет его подвижных форм. Кроме того, из приведенных выше данных следует, что синтетические лиганды, формирующие высокостабильные комплексы, могут конкурировать с фитосидерофорами за катионы железа.
Для оптимизации корневого питания растений железом в карбонатных почвах определенный интерес представляют комплексоны ароматического ряда. Специфика их структуры — ароматического кольца, заключается в увеличенной по сравнению с алифатическими комплексонами жесткости системы, препятствующей свободной ориентации атомов и функциональных групп в пространстве. Утрачивая жесткость, молекулы комплексона приобретают пространственную конфигурацию, адекватную стереохимии определенных катионов, что значительно повышает селективность комплексооб- разования. Наиболее перспективными для корневого питания растений железом оказываются производные бензольного ряда, в составе которых сочетаются два ароматических кольца: этилендиамин-N, N′-ди(2-гидроксифенил)-N, N′-диуксусная (ЭДДГА) и этилендиамин-N, N′-ди(2-гидроксифенил)уксусная (ЭДДОФА) кислоты:

Сложная стереохимия этих лигандов обусловливает напряженность образуемых хелатных циклов и определяет специфику ком- плексообразования. За исключением трехвалентного железа и редкоземельных элементов, большинство катионов не в состоянии реализовать максимальную дентатность перечисленных хелаторов. Производные бензольного ряда характеризуются также способностью образовывать высокоустойчивые комплексы с трехвалентным железом: для ЭДДОФА lg К = 20,2, для ЭДДГА lg К = 33,91. В различных почвенных условиях при адекватном подборе концентраций комплексы железа с производной гидроксибензиламинов — ЭДДГА, отличаются высокой антихлорозной эффективностью. На основании анализа диаграмм стабильности можно заключить, что в отличие от алифатических высокодентатных комплексонов ассоциация железа с ЭДДГА обеспечивает 100%-ную селективность к этому металлу даже при повышенных концентрациях кальция в среде и ее щелочной реакции (рис. 5.7).
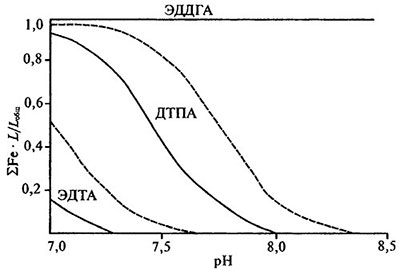
Рис. 5.7. Диаграммы стабильности для Fe-ЭДДГА, Fe-ДТПА, Fe-ЭДТА при концентрации кальция 5•10-3 моль/л [по: Lucena et al.,1988].
Сплошная линия — Fe(OH)3 аморфный, штриховая — Fe(OH)3 почвы.
Однако чрезмерно высокая стабильность хелатов ароматического ряда может негативно сказываться на процессе питания растений железом. Так, повышенная селективность N, N′-ди-(2-гидроксибензил)-этилендиамин - N, N′-диуксусной кислоты (ГБЭД)
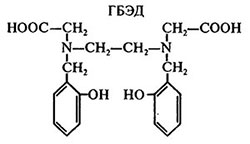
к Fe (III) и устойчивость его Fe (ІІІ)-комплексов (lg К= 39,68) препятствуют нормальному усвоению железа корнями растений (кукуруза).
Аналогичные данные в условиях гидропоники получены и для ЭДДГА. Негативное свойство последней состоит в ее участии в катализе окисления двухвалентных форм марганца и кобальта до трехвалентных форм. Несмотря на ярко выраженные свойства Fe-бензольного ряда, практическое применение этих комплексонов ограничено вследствие их высокой себестоимости.
Поглощение комплексов микроэлементов
Представления о механизмах поглощения корнем комплексных форм микроэлементов противоречивы. С одной стороны, допускается возможность непосредственного поступления экзогенных органических лигандов, не являющихся фитосидерофорами, в растительные ткани. Такие предположения основываются на результатах экспериментов с природными и искусственными хелаторами, содержащими радиоактивную метку. Например, в одно растение фасоли или подсолнечника может поступать от 9 до 40 мг углерода фракций фульвокислот (ФК). Скорость переноса углерода через корни растений возрастает с уменьшением молекулярной массы ФК. В опытах с двойной меткой (54Мn, 14С) установлено, что марганец поступает в клетки корня кукурузы в связанной форме, если фракции ФК относительно высокомолекулярны (> 10 кД), и в виде иона, если они низкомолекулярны (~380 Д). Судя по данным спектрофотометрических исследований, при корневом питании поступление в растение синтетических комплексов (Fe-ДТПА, Fe-ЭДДОФА) происходит без их разрушения.
С другой стороны, существует мнение, что в недиссоциированном виде экзогенные хелаты поступают в надземные органы растений в незначительном количестве или не поступают вовсе. В этом случае предполагается, что экзогенные хелаторы в транспорте микроэлементов по растению не участвуют, а роль лигандов сводится к обеспечению растворимости соединений микроэлементов в среде. Тогда поступление в побег экзогенного хелата в интактной форме зависит от его концентрации в среде. В экспериментах с комплексом Fe-ЭДДГА красного цвета определили его пороговую концентрацию в питательном растворе, выше которой происходила аккумуляция Fe-хелата в побеге. Если концентрация Fe-ЭДДГА была ниже порогового уровня, то в листьях содержалось лишь незначительное количество этого комплекса. Возможно, поглощение комплексных форм металлов осуществляется с помощью двух механизмов, функционирующих при высоких и низких концентрациях хелатов в среде.
Теоретический и практический интерес представляет рассмотрение вероятных механизмов поглощения высшим растением сидерофорных Fe-комплексов, продуцируемых микроорганизмами. Растениями стратегии I (см. гл. 6) эти соединения могут поглощаться как в интактном, так и в диссоциированном виде. В опытах с хлопчатником наблюдали предварительный распад сидерофора ферриоксамина В, после которого железо поглощалось растениям в нехелатированной форме. Растения огурца, однако, способны частично поглощать железо в виде интактного сидерофорного комплекса. Изученные к настоящему времени сидерофоры отличаются низкой растворимостью в липидной фазе. Поэтому в качестве вероятного механизма их транспорта через мембраны рассматривают не пассивную, а облегченную диффузию.
Один из механизмов неспецифического восстановления растением сидерофорных комплексов железа связывают с работой редуктаз. Сидерофорредуктазная активность обнаружена для нитратредуктазы. Поскольку этот фермент локализован в цитоплазме, он может участвовать в восстановлении Fe (ІІІ)-сидерофорных комплексов только после их поступления внутрь клетки. Использовать Fe (ІІІ)-сидерофоры в качестве акцептора электронов могут также НАД(Ф)Н-оксидоредуктазы, локализованные в плазмалемме клеток корня. Следовательно, растения утилизируют Fe-сидерофоры как на поверхности клеток, так и в цитоплазме.
Высокоспециализированная поглотительная система растений стратегии II, продуцирующих в ответ на недостаток железа собственные фитосидерофоры (см. гл. 6), не участвует в непосредственном поглощении сидерофорных комплексов бактериального происхождения. В этом случае ассимиляция железа растениями осуществляется непрямым способом, при котором происходит обмен лигандами между Fe-комплексами сидерофоров и фитосидерофоров. В итоге железо поглощается корнями растений стратегии II в форме Fe-фитосидерофора. Благодаря такому механизму растения стратегии II используют феррисидерофоры в качестве эффективного источника железа.
После поглощения корнем многие микроэлементы транспортируются по клеткам коры к сосудам ксилемы и по ксилеме в побег главным образом в виде хелатных комплексов. Их состав и устойчивость зависят от вида как металла, так и лиганда. С помощью электрофореза на бумаге и радиоавтографии установлено, что стабильные комплексы с соединениями ксилемного экссудата способны образовывать цинк, марганец, никель, кадмий. Железо транспортируется в побег в анионной форме в виде Fe (ІІІ)-цитрата. С компонентами ксилемного сока марганец, кальций, магний формируют более лабильные комплексы, чем цинк, медь, железо. При транспортировке в растении цинк и медь, как правило, хелатируются органическими кислотами (лимонной и яблочной), но в меньшем количестве, чем железо. Сродство микроэлементов к природным хелаторам варьирует в зависимости от природы элемента. Например, медь характеризуется большим сродством к аминокислотам, чем к органическим кислотам.
Образование комплексов микроэлементов с природными лигандами зависит от pH среды ксилемного сока, а также концентрации компонентов хелата и окружающих его соединений. В кислом диапазоне pH цинк слабо связывается аминокислотами. С повышением pH этот металл конкурирует за связи с органическими кислотами и с аминокислотами. С увеличением концентрации природных органических кислот стабильность комплексов микроэлементов в электрическом поле повышается. Высокое содержание в экссудате растений кальция препятствует комплексообразованию микроэлементов, а фосфора — формированию комплексов железа с цитратом. При избыточном содержании в растениях одних микроэлементов может изменяться состав хелатов других. Так, с увеличением концентрации цинка до токсичного уровня снижается содержание аспарагиновых комплексов меди и повышается — гистидиновых.
5.2.2. Побег
Рассмотрение роли комплексонов в усвоении микроэлементов листьями представляет теоретический и практический интерес. Освещение этого вопроса позволяет лучше понять механизмы утилизации в побеге комплексных соединений микроэлементов. Кроме того, эти сведения важны для эффективного проведения некорневых подкормок растений микроэлементами. Указанный способ получил очень широкое распространение в мировой практике растениеводства, так как позволяет оперативно, с минимальными затратами химикатов корректировать дефицит микроэлементов у культурных растений.
Комплексы железа
В процессе метаболизации побегом Fe-хелатов важная роль принадлежит фотохимическим бкислительно-восстановительным реакциям в системе Fe (III) — лиганд, сопровождающимся образованием Fe (II). Функционирование этого механизма обусловлено необходимостью восстанавливать окисленные формы железа, которые образуются при передвижении из прото- в метаксилему поглощенных корнями форм Fe (II). Для осуществления метаболических процессов в растениях нужна двухвалентная форма железа. Ключевое значение Fe (II) определяется, с одной стороны, лучшей по сравнению с Fe (III) растворимостью его минеральных солей, а с другой — меньшей устойчивостью Fe (ІІ)-хелатов, что облегчает высвобождение железа из транспортных форм и его дальнейшее включение в обмен веществ.
Фоторедукция Fe (III) до Fe (II) происходит под действием оптического излучения с длинами волн меньше 420 нм. Оптическое излучение с распределением длин волн в диапазоне 350-700 нм способно проникать-через интактный зеленый лист и вызывать фоторедукцию Fe (III) в питательном растворе. Этот процесс происходит как при прямом облучении растворов, так и при их освещении через пораженные хлорозом листья. При исключении из состава оптического облучения ультрафиолетовой (УФ) части фоторедукция Fe (III) подавляется. Стимулирующее влияние УФ-излучения на восстановление Fe (III) и синтез хлорофилла в листьях обнаружено в опытах с интактными растениями сои. При этом вызванные УФ-лучами физиологические эффекты усиливались по мере обострения Fe-дефицита и в наибольшей степени проявлялись у Fe-неэффективного сорта.
Фоторедукция Fe (III) происходит интенсивнее при участии органических соединений, в частности кислот. У растений под воздействием яркого солнечного света в аэробных условиях цитратный Fe (ІІІ)-комплекс (одна из основных транспортных форм железа в растениях) легко разлагается с образованием Fe (II). Механизм фотолиза карбоксилсодержащих хелатов железа и других микроэлементов (Мn, Со) заключается в элиминировании молекулы С02 и последовательном сокращении хелатного цикла. В процессе фотолиза образуются промежуточные продукты со связями металл—углерод. Эти продукты термически нестабильны и образованием ионов металла, а также радикалов лиганда. Фоторазложение, например, лимонной кислоты идет постепенно, с образованием многочисленных промежуточных соединений, конечные продукты — ацетон и углекислый газ. При фотораспаде ЭДТА идентифицированы следующие продукты: углекислый газ, формальдегид, ионы этилендиаминтриацетата, этилендиаминдиацегата. Фотохимические окислительно-восстановительные реакции обнаружены не только в цитратных, но и в тартратных системах. Фотохимическая активность Fe-хелатов и их физиологическое действие при некорневом питании зависят от многих факторов: природы комплексона, интенсивности и спектрального состава оптического излучения, реакции среды, содержания в составе Fe-хелата других микроэлементов.
Природа комплексона. Один из основных факторов, влияющих на фотолиз хелатов. По стимулирующему действию на фоторедукцию Fe (III) в растворе природные органические кислоты располагаются в ряду: винная > щавелевая > лимонная > яблочная > аконитовая > фумаровая. Фотохимическая активность синтетических Fe-хелатов также варьирует в зависимости от природы лиганда (табл. 5.3). При этом максимальными квантовыми выходами фотохимических реакций характеризуются карбоновые, а минимальными фосфоновые Fe-хелаты.
В ряду карбоновых комплексов также выявлены различия по этому показателю. Квантовые выходы фотохимических реакций у среднеустойчивых хелатов — Fe-ЭДЦА, Fe-HTA, выше, чем у высокоустойчивых — Fe-ДТПА, Fe-ЭДТА. Однако антихлорозное действие последних эффективнее, чем среднеустойчивых Fe-хелатов и тем более фосфоновых комплексов, которые не влияют на синтез хлорофилла в Fe-дефицитных листьях (рис. 5.8).
| Лиганд | Форма железа | |
| Fe2+ | Fe3+ | |
| НТА | 0,111 | 0,088 |
| ЭДДА | Не опр. | 0,160 |
| ЭДТА | 0,061 | 0,057 |
| ДТПА | 0,030 | 0,024 |
| ОЭДФ | 0,008 | 0,008 |
| НТФ | 0,005 | 0,005 |
| ЭДТФ | 0,006 | 0,006 |
| НСР05 | 0,002 | 0,002 |
Прим. CFc = 1 • 10-4моль/л; Fe : L — 1:1 ; pH 6,0; λобл. — 254 нм.
Очевидно, при нанесении на поверхность листа фосфоновых Fe-хелатов их фотохимическая инертность препятствует образованию биологически активных форм Fe (II). В свою очередь, различия в физиологическом действии между фотохимически активными группами средне- и высокоустойчивых Fe-хелатов могут быть обусловлены неодинаковым периодом их фотораспада. Комплексы с повышенной фотохимической активностью быстро разрушаются с увеличением продолжительности экспозиции (рис. 5.9), что сопровождается ингибированием фотовосстановления Fe (III) до Fe (II).
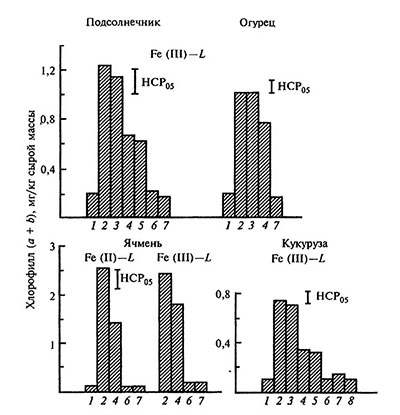
Рис. 5.8. Сожержание хлорофила в листьях при некорневом питании растений Fe-хелатами (CFe=2•10-3 моль/л; Fe L=1:1, pH 6,0) [по: Битюцкий, 1995]
Хлороз вызывали исключениями железа из состава раствора Кнопа 1 — H2O (контроль), 2 — Fe-ДТПА, 3 — Fe-ЭДТА, 4 — Fe-НТА, 5 — Fe-ЭДДА, 6 — Fe-НТФ, 7 — Fe-ОЭДФ, 8 — Fe-ЭДТФ
Следовательно, при некорневых подкормках быстрая фотодеструкция Fe-хелатов предопределяет ускоренную иммобилизацию железа в капле раствора вследствие плохой растворимости в воде его соединений. Этот процесс усугубляется относительно высокими значениями pH рабочих растворов и небольшим количеством хелатирующего агента, которое ограничено объемом капли на поверхности листа. Лучшим антихлорозным действием обладают комплексы железа, способные к интенсивной, но пролонгированной фотохимической деструкции.
Оптическое излучение. Атрибутом любой фотохимической реакции выступает оптическое излучение определенного состава и интенсивности. Наибольший фотохимический эффект достигается при действии на хелатные комплексы микроэлементов излучением в области полосы максимального поглощения. Ярко выраженные симметричные полосы поглощения при длине волны 260 нм установлены в спектрах комплексов железа с ДТПА, ЭДТА, ЭДТФ. В спектрах среднеустойчивых комплексов (Fe-ЭДДА, Fe-HTA) при той же длине волны обнаруживается плечо. Валентность железа и реакция среды изменяют оптическую плотность, но не влияют на положение максимумов поглощения в спектре Fe-комплексонатов.
По мере исключения коротковолновой области из спектра источника облучения фотохимическая активность комплексонатов железа заметно снижается. В ряду карбоновых кислот фоторедукция Fe (III) в наибольшей степени подавляется в растворах с высокоустойчивыми синтетическими комплексами (Fe-ДТПА, Fe-ЭДТА), а в наименьшей степени — с природными органическими кислотами (цитрат, малат, тартрат). Однако фотовосстановление Fe (III) в растворах с карбоновыми лигандами, в отличие от растворов фосфорсодержащих комплексонов, отмечается даже при исключении УФ-области из спектра источника облучения. Ограничение светового потока в период проведения некорневых подкормок карбоновыми Fe-хелатами нарушает усвоение листьями железа из комплекса со средней фотохимической активностью (Fe-ДТПА) и не влияет на нее, если используются фотохимически высокоактивный комплекс (Fe-HTA) или минеральные соли.
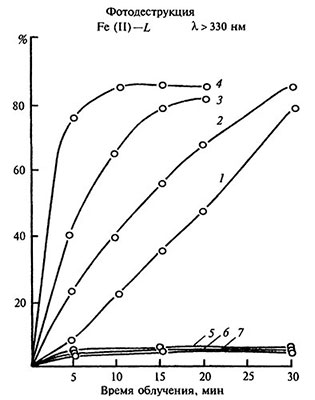
Рис. 5.9. Влияние хелатирующих агентов на фотодеструкцию Fe(II)-хелатов в водных растворах [модифицировано по: Битюцкий, 1995].
CFe=110-1 моль/л; Fe:L=1:1; pH 6,0: 1 — Fe-ДТПА, 2 — Fe-ЭДТА, 3 — Fe-НТА, 4 — Fe-ЭДДА, 5 — Fe-ЭДТФ, 6 — Fe-НТФ, 7 — Fe-ОЭДФ.
Реакция среды. В водных растворах фотохимически активных Fe-хелатов величины квантовых выходов фотохимических реакций выше в кислой среде, чем в нейтральной и щелочной. Антихлорозное действие хелатов этой группы также максимально в кислой среде. Как фотохимическая, так и физиологическая активность фосфорсодержащих комплексонатов железа сохраняется на низком уровне в широком диапазоне pH (4-9). Очевидно, пониженная скорость фотообразования Fe (II) лимитирует усвоение железа листьями растений при нейтральной и щелочной реакциях среды. Отметим, что эти результаты получены в экспериментах с гомогенными растворами Fe-комплексонатов. По этой причине отрицательное влияние повышенных значений pH на антихлорозную активность Fe-комплексов при их нанесении на лист не может быть интерпретировано как деструкция хелатов.
Влияние микроэлементов. Клеточный сок растений представлявет собой многокомпонентную систему, в которой железо находится в окружении различных соединений других микроэлементов. Многие из металлов-микроэлементов участвуют в регуляции фотохимической и физиологической активности Fe-хелатов. Воздействие на фотовосстановление Fe (III) микроэлементов определяется их природой и концентрацией, видом органического лиганда и величиной соотношения железо : микроэлемент: лиганд в растворе (рис. 5.10). При соотношении Fe: L = 1:2, достаточном для хелатирования всех металлов композиции, обнаружено сильное (особеннo на начальных этапах облучения) ингибирование фотохимического восстановления Fe (III) в присутствии меди и марганца. Например, при эквимолярном соотношении этих металлов с Fe (III) скоростъ его фотовосстановления уменьшается по сравнению с контролем на 50-80%. По сравнению с медью и марганцем, влияние цинка на фотохимические реакции железа меньше. При эквимолярном соотношении Fe : L, недостаточном для хелатирования привносимых микроэлеменов, не происходит подавления фотовосстановления Fe (III) ни марганцем, ни тем более цинком. В отличие от этого прибавление к эквимолярным комплексам железа меди может индуцировать, в силу ее высокой комплексообразующей способности, разрушение Fe-хелатов и образование нерастворимых в воде гидроксидов железа.
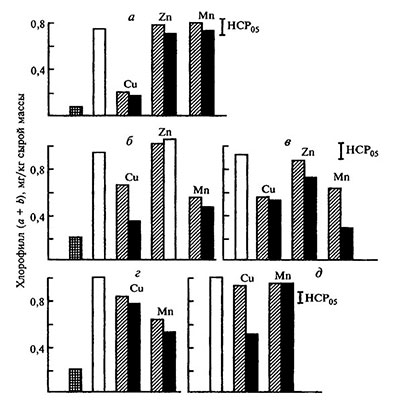
Рис. 5.10. Фотохимическое восстановление хелатированного Fe(III) до Fe(II) в водных растворах [по. Битюцкий, 1995].
CFe=1:10-3 моль/л, рН 5,5, λобл=254 нм, экспозицию — 3 и 6 мин
а — Fe(III)•ДТПА=1:2, t=3 мин; t=3 мин; б — то же, t=6 мин; в — Fe(III)•ЭДТА=1:2, t=6 мин, г — Fe(III) ЭДТА=1:1, t=6 мин. Светлые столбики — источники железа без микроэлементов, заштрихованные — FeL:M = 1:0,5, темные — FeL:M=1:1
Фотохимические эффекты микроэлементов аналогичны физиологическим (рис. 5.11). Так, действие на хлорозне листья огурца хелатов железа с соотношением Fe : L = 1 :2 сильно нарушается, если в растворах содержатся медь и марганец, а цинк на него не влияет. В то же время излечивание хлороза не тормозится при использовании тех же количеств марганца из смеси с эквимолярным (Fe : L = 1:1) комплексом железа (Fe-ЭДТА) и из FeS04. Обработка листьев смесями этих соединений железа с медью подавляла его антихлорозное действие. В данном случае негативный физиологический эффект обусловлен потерей водорастворимости железа при взаимодействии с медью, о чем свидетельствует появление осадка.
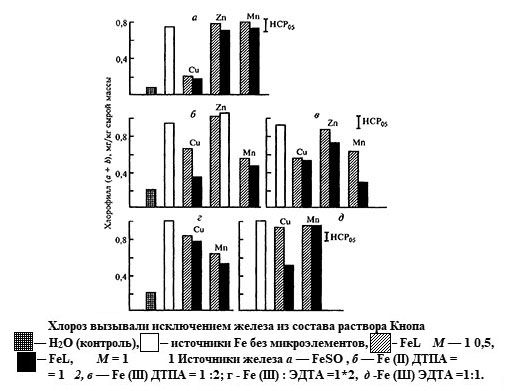
Рис 5. 11. Эфективность некорневого питания огурца соединениями железа CFe=2×10-3 моль/л pH 5,5) [по: Битюцкий, 1995]
Ингибирование металлами фотохимического восстановления Fe (III) обусловлено известным в фотохимии эффектом тушения электронно-возбужденных состояний молекул. Существует точка зрения, что если тяжелый атом посторонний (нецентральный), то достоверного тушения не происходит в результате более эффективного влияния центрального иона комплекса на спин-орбитальное взаимодействие. Видимо, по этой причине марганец в композиции с эквимолярным комплексом Fe (III) не оказывает существенного воздействия на его фотовосстановление и физиологическую активность.
Результаты исследований с композициями микроэлементов позволяют объяснить один из возможных механизмов нарушения обмена железа у растений в условиях его недостатка. Вероятна следующая последовательность событий. Недостаток железа в питательной среде индуцирует сильное накопление в корнях и отдельных органах побега многих видов растений органических кислот и металлов-микроэлементов, особенно марганца (табл. 5.4, 5.5). Избыток хелатирующих соединений способствует ингибированию (тушению) микроэлементами фотохимических реакций в системе железо — хелатирующим агент, что усугубляет дефицит функционально активных форм Fe (II) и в целом негативно сказывается на фитометаболизме.
| Часть растения | Железо | Марганец | Цинк | |||
| Контроль | HCO3- | Контроль | HCO3- | Контроль | HCO3- | |
| Листья: верхние | 96 | 37 | 92 | 126 | 85 | 78 |
| средние | 78 | 49 | 104 | 110 | 82 | 65 |
| нижние | 70 | 58 | 83 | 81 | 73 | 69 |
| Стебель | 31 | 18 | 46 | 53 | 80 | 30 |
| Корень | 613 | 172 | 47 | 111 | 75 | 130 |
В естественных условиях фоторазложение органических кислот, сопровождающееся восстановлением Fe (III) до Fe (II), локализовано в листе и представляет собой один из механизмов адаптации растений к Fe-стрессу. Физиологическое значение фоторедукции Fe (III) возрастает с обострением Fe-дефицита и находит наибольшее выражение у Fe-неэффективных видов. Вместе с тем образование двухвалентной формы хелатированного железа нельзя считать достаточным условием для его включения в метаболизм побега. Этот процесс, вероятно, происходит только после фотодеструкции Fe-xeлата. Повышенные концентрации в тканях микроэлементов и высокие значения реакции среды негативно отражаются как на фотохимическом восстановлении железа в листе, так и на функционировании Fe-редукгаз корня и рассматриваются в качестве физиологических факторов обострения карбонатного хлороза растений.
| Органические кислоты | Fe, 11 мкмоль/л,
(контроль) |
Fe, 11 мкмоль/л, + HCO3- | Fe, 4 мкмоль/л, + HCO3- |
| Цитрат | 9,95 | 14,59 | 17,21 |
| Малат | 4,09 | 4,86 | 5,81 |
| Малонат | 5,74 | 6,65 | 7,60 |
| Аконитат | 0,13 | 0,15 | 0,14 |
| Сукцинат | 111,62 | 76,02 | 87,74 |
| Фумарат | 7,98 | 6,64 | 6,32 |
Комплексы других микроэлементов
Представления о необходимости хелатирования марганца, цинка, меди и кобальта, в отличие от железа, при проведении некорневых подкормок растений противоречивы, а сведения по этому вопросу малочисленны. Возможно, преимущества хелатов проявляются при транслокации этих микроэлементов из обработанных листьев к другим органам растений. Однако для многих минеральных солей металлов характерна довольно высокая миграционная способность. В опытах с карликовым горохом установлено, что усиление передвижения микроэлементов по растению под действием комплексонов происходит не всегда. Специфика этого явления обусловлена природой металла и органического лиганда. В основном соли 3d-элементов растворимы в воде в более широком диапазоне pH, чем соли железа, а это существенно снижает эффективность действия комплексонов как на стадии сорбции металлов кутикулой, так и на последующих этапах их метаболизма. В ряде случаев отмечают подавление синтетическими хелаторами эффектов, вызываемых минеральной формой микроэлемента, что свидетельствует о снижении интенсивности его включения в метаболизм растений из органических комплексов. Подавление физиологической активности микроэлементов при их нанесении на поверхность листа наиболее четко выражено у фосфорсодержащих комплексонов. В некоторых случаях такое ингибирование может быть обусловлено изначально слабой растворимостью в воде фосфоновых комплексов. Кроме того, потеря растворимости соединений микроэлементов вероятна и при фотолизе фосфоновых комплексов, который может сопровождаться образованием неорганических фосфатов.