

В этой главе содержится информация о специфической и неспецифической адаптации высших растений к недостатку микроэлементов в среде. Рассмотрены современные представления о механизмах специфической адаптации к условиям Fe-cmpecca. Приведена подробная характеристика групп растений, различающихся по механизмам адаптации к недостатку железа (стратегия I и стратегия II).
6.1. Неспецифическая адаптация
Условно возможные механизмы адаптации растений к недостатку питательных элементов подразделяют на неспецифические и специфические. Действие неспецифических механизмов не связано с прямой реакцией растений на недостаток конкретного элемента. В этом случае мобилизация микроэлементов в почве и улучшение минерального питания растений достигаются за счет комплекса физико-химических и биохимических процессов в ризосфере корней. Ключевая роль в функционировании неспецифических механизмов принадлежит корневым выделениям, в частности корневым слизям. При специфической адаптации у растений отмечается прямая физиологическая реакция на дефицит конкретного элемента.
Корневые слизи. На поверхности корня находится слой ризодермальных клеток, связанных с ризосферой тонкой оболочкой слизи. В выделении слизей участвует внешний слой эпидермиса. Особенно много слизей выделяют апикальные зоны корней. Синтез растительных слизей в клетке и их перемещение из протопласта к плазмалемме осуществляются комплексом Гольджи, на поверхность корня слизь выделяется через клеточные стенки. Слизи представляют собой высокомолекулярный гелевый материал фибриллярной структуры. Корневые слизи состоят в основном из полисахаридов, включающих в зависимости от вида растений от 20 до 50% полиуроновых кислот. Сеть фибрилл формирует поровую систему, которая вместе с апопластом образует свободное пространство (СП) корня. Взаимодействие мономеров фибрилл осуществляется через солевые мостики между карбоксильными группами. Эта структура стабилизирована водородными связями ОН-групп. Важную роль в формировании солевых мостиков играют ионы кальция. С повышением в среде концентрации ионов кальция степень гидратированное фибриллярной сета возрастает. Слизи играют важную роль в защите апикальных зон корня при его передвижении в почве и в поглощении ионов. Будучи высоко гидратированными, слизи непосредственно контактируют с поверхностью почвы и усиливают обменные процессы корня.
Взаимодействие кальция корневых слизей с ионами среды может быть обменным и необменным. Обменное взаимодействие характерно для химических форм, электростатически связанных с сетчатыми структурами. В результате обменных реакций и пассивного транспорта в слизях поглощаемые ионы способны мобилизоваться в апопласте клеток корня. По типу обменного происходит взаимодействие корневых слизей с Са2+, Mg2+, Fe2+, Mn2+. Необменное взаимодействие отмечается в том случае, если формы ковалентно связаны с участками фибриллярной структуры. Такие химические формы не могут быть перенесены к плазмалемме, они снижают гидратированность фибриллярной сети и ее пористость.
Необменное взаимодействие характерно для алюминия и многих тяжелых металлов (особенно для меди) при их высоком содержании в почве. В нормальных условиях, т. е. при относительно низком содержании в среде микроэлементов и высоком содержании оснований корневые слизи, сохраняя гидратированность и объем, способны участвовать в поглощении элементов питания. При недостатке влаги в почве выделение корневых слизей в ответ на возрастающее механическое сопротивление корню усиливается, что положительно сказывается на поступлении в растения микроэлементов. Благоприятное косвенное влияние на этот процесс оказывает также увеличение корневыми слизями в почве количества водопрочных агрегатов.
Низкомолекулярные корневые выделения. Ризосфера представляет собой область почвы (среды), в которой концентрируются разнообразные по химическому составу относительно низкомолекулярные корневые выделения. Их главными компонентами являются сахара, органические кислоты, аминокислоты и фенолы, обычно превалируют сахара и органические кислоты. Общее количество и соотношение компонентов низкомолекулярных корневых выделений сильно варьируют в зависимости от вида растений и их минерального статуса. В нестерильных условиях точное определение количества низкомолекулярных соединений затруднено вследствие их быстрой утилизации микроорганизмами.
Наиболее сильное прямое действие на мобилизацию микроэлементов в почве оказывают органические кислоты, аминокислоты и фенолы. Воздействие на этот процесс сахаров обычно незначительно. Повышение растворимости соединений микроэлементов под влиянием корневых выделений происходит за счет восстановления окисленных форм микроэлементов (например, Мn) и образования растворимых хелатов (Mn, Fe, Zn) (рис. 6.1). Специфическая роль фенолов и органических кислот в мобилизации соединений железа будет рассмотрена ниже.
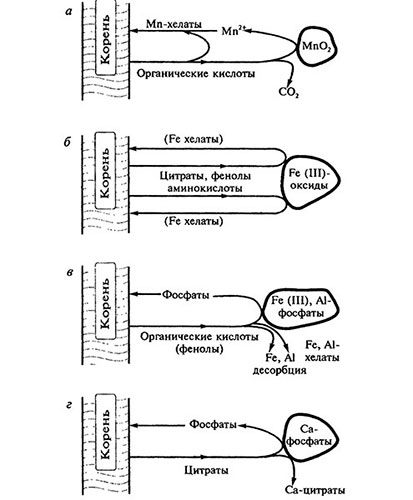
Рис. 6.1. Растворение корневым выделениями плохо растворимых неорганических соединений в ризосфере растений [модифицировано по: Marschner, 1997; слюбезного разрешения Academic Press Limited]
Для мобилизации микроэлементов почвы важен ацидофицирующий эффект корневых выделений. Здесь не рассматриваются механизмы ацидофикации, связанные с прямой реакцией растений на дефицит микроэлементов. Значение pH почвы, не затронутой деятельностью корней, может превышать величину pH ризосферы более чем на две единицы. В пределах индивидуальной корневой системы разница значений pH вдоль оси корня и между основными и боковыми корнями также может достигать двух единиц. Как правило, у растений, произрастающих на кислых почвах, величина pH выше у корневого апекса и ниже в субапикальной зоне.
Изменения реакции среды в ризосфере связаны с нетто-выделением корнями Н+, НС03- или ОН- и органических кислот. Кроме того, органические кислоты появляются в ризосфере вследствие активности микроорганизмов, стимулируемой выделяемыми корнями органическими соединениями. Значительное влияние на pH ризосферы оказывает баланс скорости поглощения корнями катионов и анионов. В случае более интенсивного поглощения корнями катионов усиливается продуцирование Н+ и pH ризосферы понижается. При более интенсивном поглощении корнями минеральных форм анионов pH ризосферы повышается за счет выделения НС03- или ОН-.
Особенно важную роль в регуляции ионного баланса растений и pH ризосферы играют источники азотного питания. Если это нитраты, то скорость выделения корнями НС03-, как правило, выше, чем протонов.
При аммонийном питании наблюдается обратная зависимость. В нейтральных и щелочных почвах подкисление ризосферы у растений, питающихся аммонийными формами азота, способствует усилению поглощения корнями бора, железа, марганца, цинка. В кислой среде понижение значений pH ризосферы у растений, выращиваемых с использованием аммонийных источников питания, может быть причиной резкого торможения роста и продуктивности растений. Этот эффект обусловлен тем, что продуцируемые растением в апопласт протоны препятствуют поступлению и оттоку в листья катионов. Устойчивая нейтрализация среды у растений на аммонийном фоне способствует устранению отмеченного выше отрицательного физиологического эффекта.
У бобовых, обеспечивающих потребности в азоте с помощью симбиотической азотфиксации, соотношение минеральные катионы/анионы и соответственно экскреция Н+ корнями выше, чем у растений тех же видов на нитратном фоне. Поэтому в тканях растений, фиксирующих азот, содержание микроэлементов выше по сравнению с растениями тех же видов, питающихся нитратными формами азота. О масштабах подкисления ризосферы бобовыми можно судить по следующему примеру. Растения люцерны, фиксирующие атмосферный азот, при урожае 10 т/га снижают кислотность почвы на величину, эквивалентную внесению в почву 600 кг/га СаС03. При длительном выращивании бобовых на одном месте, а также в севооборотах с высокой долей бобовых ацидофикация почвы растениями, фиксирующими азот, может способствовать повышению концентрации марганца до токсичного уровня.
На подвижность микроэлементов в почве оказывает влияние и величина окислительно-восстановительного потенциала ризосферы, которая зачастую существенно отличается от этого показателя для почвы, не затронутой функционированием корней. Зона действия корней на величину Eh может распространяться на несколько миллиметров от поверхности ризопланы. Кроме того, Eh ризосферы изменяется под влиянием функционирования микроорганизмов, концентрация которых в ризосфере существенно выше, чем за ее пределами. Как уже упоминалось (см. 2. 2. 2), уменьшение Eh коррелирует с повышением растворимости многих микроэлементов.
Выделение корневых экссудатов тесно связано с минеральным статусом растений. В условиях недостатка элементов питания (не обязательно микроэлементов) количество низкомолекулярных корневых выделений увеличивается. Это эффект установлен для растений различных видов при недостатке в почве, например, калия и фосфора. К тому же нередко изменяется и качественный состав корневых выделений. Так, в условиях К-дефицита у кукурузы соотношение в корневых выделениях сахаров и органических кислот изменяется в пользу кислот. У люцерны и карликового гороха при недостатке фосфора усиливается выделение лимонной кислоты, а у арахиса фумаровой.
Индуцированное недостатком макроэлементов выделение корневых экскретов может благоприятно сказываться и на доступности микроэлементов для растений. Высокая скорость локальной экссудации корнями органических кислот и протонов обеспечивает растениям экологическое преимущество, особенно в почвах с большой буферной емкостью. Этот принцип довольно хорошо реализован у растений (белый люпин) с протеоидной (кластерной) корневой системой. Высокая плотность корней в кластере способствует более эффективному подкислению почвы и лучшей адаптации растений к условиям недостаточной подвижности в среде элементов питания. Функционирование протеоидной корневой системы белого люпина обеспечивает локальное подкисление даже карбонатных почв. При нормальном снабжении люпина растворимыми фосфатами подавляется формирование протеоидных корней и выделение ими лимонной кислоты.
В целом количество корневых выделений в ризосфере зависит от множества эндогенных и экзогенных факторов. К важнейшим из них относятся биологические особенности культур, динамика питательных веществ в ризосфере, подвижность минеральных элементов, минеральный статус растений. Кроме того, на выделение корневых экссудатов оказывает влияние механическое сопротивление почвы. С повышением плотности почвы количество корневых выделений в ризосфере возрастает. Вероятно, с увеличением механического сопротивления тормозится в основном рост корня, а транспорт к нему продуктов фотосинтеза не меняется. В результате потребление продуктов фотосинтеза в расчете на единицу длины корня может возрастать в несколько раз. Продуцирование корневых выделений иногда оказывает положительное влияние на питание растений микроэлементами. Однако рассмотренные здесь физиологические реакции растений, индуцированные различными факторами, напрямую не связаны с ответом растительного организма на дефицит микроэлементов в почве.
6.2. Специфическая адаптация
Как отмечалось выше, специфические механизмы адаптации обеспечивают прямую физиологическую реакцию растений на дефицит микроэлементов, направленную на повышение их подвижности и доступности в среде. К настоящему времени наиболее детально изучена реакция растений на дефицит железа.
Эта реакция обусловлена комплексом адаптивных механизмов. Химические способы повышения растворимости и доступности железа растениям ограничиваются восстановлением Fe (III) до Fe (II), уменьшением pH ризосферы и продуцированием растением хелатирующих соединений. Активность этих процессов зависит как от специфики генотипа растения, так и от его обеспеченности железом.
Генотипически обусловленные различные адаптивные реакции растений на дефицит железа классифицируют как стратегию I и стратегию II. Сначала хемотаксономическую границу двух стратегий проводили между одно- и двудольными растениями. В настоящее время группа стратегии I объединяет все двудольные и однодольные растения, не относящиеся к семейству злаков (Роасеае). Соответственно к группе стратегии II принадлежат только растения семейства злаков.
6.2.1. Растения стратегии I
Механизмы адаптации
Растения этой группы характеризуются по крайней мере двумя четкими ответными реакциями на Fe-дефицит (рис. 6. 2): активацией в клетках корней редуктаз, участвующих в восстановлении Fe (III) до Fe (II) и повышением нетто-экскреции корнями протонов. Во многих случаях эти реакции сопровождаются продуцированием корнями редуцирующих или хелатирующих соединений, например фенолов.
Наиболее чувствительная к Fe-дефициту и типичная реакция растений стратегии I заключается в увеличении активности редуктаз. В условиях достаточной обеспеченности железом и его дефицита редуктазы растений имеют сходные характеристики и локализованы в плазмалемме ризодермальных клеток. При дефиците железа увеличение редуктазной активности корней сопровождается появлением в плазмалемме нового белка с молекулярной массой около 70 кД. Донорами электронов для Fe-редуктазы служат НАДФН или НАДН. Возрастание редуктазной активности ведет к интенсивному восстановлению растворимых Fe (III)-хелатов и повышению скорости их расщепления, которое, по-видимому, необходимо для поглощения железа корнями. До сих пор не раскрыт механизм транспорта восстановленного железа в цитоплазму. Пока не ясно, участвует в этом процессе редуктаза или какой-то другой белок. Известно, однако, что активность редуктаз и скорость поглощения железа находятся под строгим генетическим контролем и, вероятно, контролируются одним доминантным геном.
Высвобождение корнями в условиях Fe-дефицита протонов связывают с функционированием в плазмалемме Н+-АТФаз. Вклад в этот процесс редуктаз, по-видимому, незначителен. Работа Н+-АТФаз сопровождается снижением величины pH в ризосфере, что способствует как увеличению растворимости неорганических соединений железа, так и оптимизации условий для работы редуктаз. Оптимальное для работы последних значение pH находится в кислом диапазоне (pH 5).
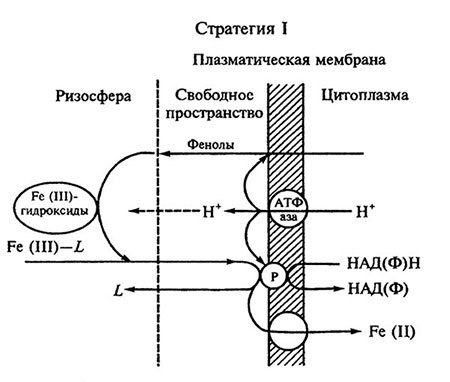
Рис 6.2. Поглощение железа корнями растений стратегии I [вопроизведено по: S. Mori. Metal lons in Biological Systems. 1998 Vol. 35 P. 217;с любезного разрешения Marcel Dekker. lnc.]
В ряде случаев первые две реакции растений стратегии I сопровождаются появлением корневых экссудатов восстанавливающих/хелатирующих веществ. В их составе идентифицированы соединения фенольной природы. Среди них важная роль в мобилизации железа принадлежит кофейной кислоте, которая может восстанавливать Fe (III) до Fe (II). Одна молекула кофейной кислоты способна восстановить девять атомов Fe (III) до Fe (II). Реакция состоит из двух основных этапов (рис. 6.3). Первый этап очень короткий. Он включает перенос пяти электронов от молекулы кофейной кислоты до Fe (III) и образование промежуточного продукта I неидентифицированной природы. На втором этапе, относительно длинном, промежуточный продукт распадается с образованием двух компонентов А и В (их природа также точно не установлена). С участием компонентов А и В осуществляется перенос еще четырех электронов на Fe (III). Рассмотренная реакция восстановления кофейной кислотой Fe (III) до Fe (II) может происходить в свободном пространстве и ризосфере корня. Вместе с тем оптимум этой реакции находится в кислой среде (pH 3-4), что ограничивает ее адаптивное значение в почвах с высокими значениями pH и большой буферной емкостью.
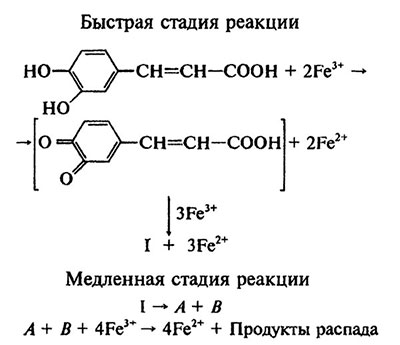
Рис. 6.3. Этапы окислительно-восстановительных реакций между Fe(III) и кофейной кислотой [по: Deiana et al.,1992]
Рассмотренные ответные реакции растений стратегии I на Fe-стресс тесно связаны с морфологическими изменениями в структуре клеток корня — образованием в его апикальной зоне ризодермальных переходных клеток. Они характеризуются интенсивным развитием протуберанцев клеточной стенки, плотной цитоплазмой с большим числом митохондрий, шероховатым эндоплазматическим ретикулумом, амебоидными пластидами.
Н+-АТФазы, активность которых индуцируется Fe-стрессом, локализованы на плазмалемме переходных клеток. В этих клетках синтезируется лимонная кислота, аккумулирующаяся в корнях в период Fe-дефицита и играющая важную роль в транспорте железа в побег. Как известно, синтез цитрата и других органических кислот происходит в митохондриях. Скопление митохондрий в переходных клетках, очевидно, можно рассматривать как приспособительный фактор, обеспечивающий растения в условиях Fe-дефицита необходимыми для растворения железа хелатирующими агентами. Кроме того, органические кислоты (малат, цитрат) могут быть источниками дополнительных протонов, выделяемых корнями. У растений томата аккумуляцию лимонной кислоты в корнях при Fe-дефиците связывают со снижением активности аконитазы. Вместе с тем в корнях бобовых активность аконитазы при Fe- дефиците не меняется, несмотря на накопление лимонной кислоты.
Следует, однако, отметить, что избыточное накопление в корне органических кислот в сочетании с аккумуляцией некоторых микроэлементов (Сu, Мn), обычно наблюдаемой в условиях Fe-дефицита, может привести к подавлению фотохимической активности хелатов, участвующих в транспорте железа в побег, и обострению признаков хлороза (подробнее см. 5. 2. 2). В целом образование переходных клеток рассматривается в настоящее время как типичный морфологический признак растений стратегии I.
В условиях Fe-дефицита дифференцировка ризодермы, ведущая к образованию в корне переходных клеток, иногда сопровождается аналогичной дифференцировкой паренхимных клеток, окружающих сосуды ксилемы. Для паренхимных клеток типичны плотная цитоплазма и большое скопление митохондрий. Возможно, эти клетки участвуют в активном транспорте железа в ксилему. Кроме того, переходные клетки могут формироваться в побеге и жилках листьев. Сигнал (вероятно, гормональной природы) для начала дифференцировки и формирования переходных клеток поступает из молодых развивающихся тканей, наиболее остро «ощущающих» недостаток железа. Например, обработка листьев абсцизовой кислотой и цитокинином увеличивает транспорт железа из корней в побег. Под влиянием ауксина, поступающего из побега в корни, усиливается работа АТФ-зависимой протонной помпы, что также облегчает транспорт железа в побег.
Для растений стратегии I разных видов обнаружена тесная положительная корреляция между активностью адаптивных реакций на Fe-дефицит (активность редуктаз и протонной помпы) и устойчивостью к заболеванию карбонатным хлорозом. Эта закономерность проявляется и у растений одного вида. При возделывании на карбонатных почвах культурные растения устойчивых генотипов отличаются высокой продуктивностью.
Влияние внешних факторов
Выявлено несколько факторов, снижающих эффективность работы механизмов стратегии I и обостряющих проявление карбонатного хлороза у растений. Наиболее важные среди них — повышенные концентрации в питательной среде бикарбонатов, нитратов, фосфатов и тяжелых металлов.
Ключевая роль в регуляции обмена железа у растений стратегии 1 принадлежит бикарбонатам, входящим как составная часть в буферную систему карбонатных почв. Под влиянием бикарбонатов (НСО3-) снижается продуктивность и нарушается обмен железа у растений стратегии I многих видов (подсолнечник, бобы, виноград). Переувлажнение и уплотнение почвы усугубляют заболевание растений хлорозом, что происходит вследствие повышения концентрации в почве бикарбонатов. Механизм физиологического действия НСO3- связывают с регуляцией pH среды. Увеличение концентрации бикарбонатов в почве способствует не только повышению pH и снижению подвижности соединений железа, но и нейтрализации продуцируемых корнями протонов и подщелачиванию ризосферы растений стратегии I. В результате нарушаются функционирование Fe-редуктаз корня и поглощение железа корнями растений этой группы. Нарушения на уровне поглощения железа корнем негативно отражаются на транспорте этого микроэлемента в побег и распределении в листьях.
Сведения о влиянии бикарбонатов на реакцию среды цитоплазмы клеток листьев противоречивы. Увеличение под воздействием НС03- значения pH зафиксировано в опытах с листьями подсолнечника и подвоев винограда. Повышением значений pH цитоплазмы клеток объясняют инактивацию железа в листьях. Однако в опытах с водными культурами подсолнечника эта точка зрения не подтверждена. Реакция растений на воздействие бикарбонат-иона биологически специфична. По мнению ряда авторов, наибольшей устойчивостью характеризуются Fe-эффективные виды.
Обострение признаков карбонатного хлороза у растений стратегии I может наблюдаться и при использовании нитратов в качестве источника азотного питания. Механизмы действия нитратов и бикарбонатов на обмен железа у растений могут быть сходны и заключаются в регуляции реакции среды. Выделение ОН- и подщелачивание цитоплазмы клеток листьев происходят в процессе восстановления нитратов на стадии протонирования нитрита. Возможно, при нитратном питании в листьях и корнях растений происходят аналогичные процессы: повышается pH апопласта, что отрицательно сказывается на восстановлении Fе(ІІІ)-соединений, в основном Fe(III)-цитрата. Поэтому транспорт железа в клетки корней и листьев уменьшается. При этом рост и формирование новых листьев (подсолнечник) более чувствительны к недостатку железа, чем содержание в листьях хлорофилла. Потребность в железе у растений подсолнечника на питательной среде с нитратом в 1,6 раза выше, чем на аммонийной среде.
В быстро растущих молодых листьях часть (около 10%) апопласта характеризуется высокими значениями pH (6,5-7). Обнаружена высокая отрицательная корреляция между pH апопласта или сока листьев и содержанием в последних хлорофилла. Одна из причин подавления роста меристем листьев состоит в уменьшении активности рибонуклеотидредуктазы. Этот фермент содержит железо и катализирует восстановление рибозы до дезоксирибозы. В результате при нарушении снабжения молодых листьев железом подавляется синтез ДНК. В наибольшей степени симптомы хлороза проявляются при содержании в питательной среде нитратов и бикарбонатов. Видовые и сортовые различия в азотном обмене и нитратредуктазной активности растений, вероятно, взаимосвязаны с генотипической спецификой их Fe-адаптивных реакций.
Мнения о характере влияния фосфатов на Fe-обмен растений неоднозначны. В полевых опытах с длительным и интенсивным применением фосфорных удобрений не выявлено корреляции между содержанием в почве фосфора и степенью проявления у растений хлороза. В водных культурах повышенная концентрация фосфора в питательном растворе может индуцировать типичные признаки хлороза растений. Одна из причин появления последнего заключается в образовании малорастворимых в воде фосфатов металла, другая состоит в подавлении активности Fe-редуктаз корня. В природных условиях этот эффект практически не наблюдается, вероятно, вследствие пониженного содержания фосфора вокруг корневых систем и пространственной разобщенности зон поглощения этих элементов на поверхности корня.
Подавление Fe-редуктазной активности корней происходит и под влиянием тяжелых металлов (медь, никель, молибден, цинк, марганец). Наибольший ингибирующий эффект вызывают медь и никель. Толерантность к действию тяжелых металлов выше у растений Fe-эффективных видов стратегии I, с большей редуктазной активностью.
6.2.2. Растения стратегии II
Механизмы адаптации
Группа стратегии II представлена только растениями семейства злаков. Для растений стратегии II характерны особые хелатирующие вещества (фитосидерофоры), выделяемые в условиях Fe-дефицита, и высокоэффективная по отношению к их комплексам с Fe (III) поглотительная система (транслокатор) (рис. 6.4). Продуцирование фитосидерофоров происходит также в ответ на дефицит цинка. Однако механизм действия цинка, в отличие от железа, на этот процесс пока не изучен. Фитосидерофоры представляют собой производные никотинаминов (мугеневая, авеневая кислоты):
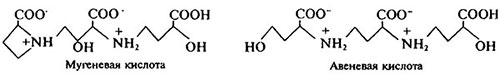
Предшественниками фитосидерофоров служат метионин и никотинамин:
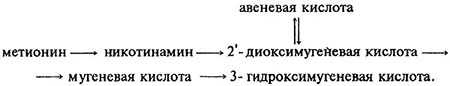
Фитосидерофоры, такие как мугеневая кислота (слева), образуют с Fe (III) высокоустойчивые комплексы, lg К порядка 33.
Транспортная система растений стратегии II локализована в плазмалемме клеток корня. Этот транслокатор обладает высокой специфичностью к Fe (ІІІ)-фитосидерофорам и в основном участвует в переносе железа. Недостаток железа не только индуцирует синтез фитосидерофоров, но и повышает скорость работы транспортирующей системы. Несмотря на то что фитосидерофоры могут образовывать комплексы с другими микроэлементами: цинком, медью, марганцем, сродство транспортной системы растений стратегии II к этим комплексам низкое. Тем не менее фитосидерофоры могут косвенно увеличивать скорость поглощения корнем таких микроэлементов за счет повышения их мобильности в ризосфере и апопласте.
Выделение фитосидерофоров корнями, индуцированное Fe-стрессом, происходит согласно довольно четкому суточному ритму. Оно начинается примерно через два часа после начала освещения и продолжается в течение четырех-шести часов. Интенсивное выделение в течение короткого периода фитосидерофоров снижает вероятность их разложения микроорганизмами ризосферы и повышает эффективность мобилизации железа в почве.
Механизмы регуляции суточной ритмики выделения фитосидерофоров полностью не ясны. Активность ферментов биосинтеза фитосидерофоров: никотинаминсинтетазы и никотинаминтрансферазы, не подвержена соответствующим колебаниям. Суточная динамика выделения фитосидерофоров коррелирует с появлением и исчезновением в клетках кончиков корней везикул и, вероятно, не связана с динамикой продуктов фотосинтеза.

Рис. 6.4. Поглощение железа корнями растений стратегии II [воспроизведено по: S. Mori. Metal lons in Biological Systems. 1998 Vol. 35 P. 218; с любезного разрешения Marcel Dekker. lnc.]
Кроме корней фитосидерофоры обнаружены в побеге, как правило, в молодых растущих тканях. Однако в наибольшей степени фитосидерофоры выделяются в апикальных зонах корней. В этих же зонах, вероятно, сосредоточен основной синтез фитосидерофоров. Во всяком случае наибольшее накопление никотинаминов также регистрируют в кончиках корней. Первоначально содержание фитосидерофоров в побеге объясняли их поступлением из апикальных частей корней. Затем была выявлена разница состава фитосидерофоров в побеге и корне. Кроме того, эти природные хелаторы не обнаружены во флоэмном соке. Поэтому побег, наряду с кончиками корней, можно рассматривать как одно из вероятных мест синтеза фитосидерофоров в растении. О физиологических функциях фитосидерофоров в листьях практически ничего не известно. С учетом свойств этих соединений допускается возможность их участия в хелатировании Fe (III) и оптимизации транспорта железа в побеге. Фитосидерофоры в побеге, как и в корнях, могут осаждаться в апопласте клеток листьев в виде комплексов с Fe (III). До сих пор, однако, неизвестно, существует ли в плазматической мембране листьев растений стратегии II специфический переносчик фитосидерофорных комплексов железа.
С помощью 59Fe у ячменя в базальной части листьев обнаружена зона меристематических клеток, которая получила название «дискриминационный центр» транспорта питательных веществ (рис. 6.5). Предположительно, этот центр участвует в контроле за перераспределением железа в растениях стратегии II.
Транспорт Fe-фитосидерофорных комплексов в побег зависит от условий освещения (рис. 6.5). Например, транспорт 59Fe из корней в полностью затемненные листья ячменя прекращается. Когда затеняли только среднюю часть листьев, поступление железа в мезофилл затемненных участков листовой пластинки уменьшалось, а транспорт 59Fe происходил в жилках листа. Следовательно, радиальный транспорт железа в листе из ксилемы к клеткам мезофилла регулируется светом. Возможно, светозависимый контроль за поступлением железа в клетки мезофилла осуществляют хлоропласты, так как оно сосредоточено в этих структурах листа. Освещение хлоропластов ячменя в течение 1 ч способствовало увеличению поглощения хлоропластами железа с 0,4 до 1,15 мкг/мг хлорофилла. Поглощение железа хлоропластами на свету почти полностью подавляется дихлорфенилдиметилмочевиной (10-5 моль/л), значит, оно не связано напрямую с фитохромной системой. Конкретные механизмы транспорта железа в хлоропласт к настоящему времени изучены недостаточно.
В темноте хлоропласты не поглощают, а напротив, выделяют железо. Этот феномен лежит в основе ремобилизации железа в листьях, более интенсивной в темноте. Затемнение нижних листьев у Fe-дефицитных бобов способствует накоплению этого микроэлемента в верхних листьях и уменьшению степени заболевания растений хлорозом. Ремобилизация железа в листьях, вероятно, осуществляется следующим образом: хлоропласт → цитоплазма → апопласт → флоэма.
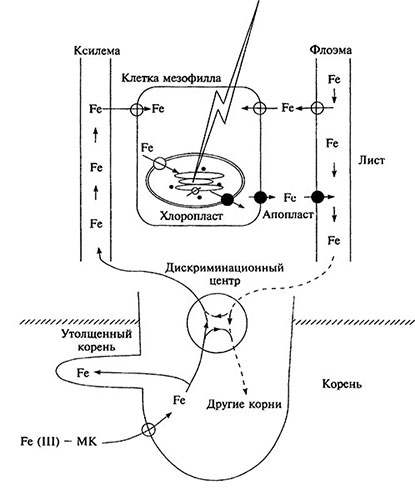
Рис. 6.5. Транспорт железа в растении ячменя (стратегия II) в темноте и на свету [воспроизведено по: S. Mori. Metal lons in Biological Systems. 1998 Vol. 35 P. 234; с любезного разрещения Marcel Dekker. lnc.]
Белыми кружками обозначены переносчики, активируемые на свету, черными - в темноте.
Устойчивость видов семейства злаков, а также отдельных форм в пределах одного вида к заболеванию карбонатным хлорозом тесно связана с их способностью выделять фитосидерофоры. По активности выделения фитосидерофоров в условиях Fe-дефицита виды злаков располагаются в ряду: пшеница > ячмень > рожь, овес > кукуруза >>> сорго. Очень мало сидерофоров выделяют корни риса при его затоплении.
Недостаток железа индуцирует синтез сидерофоров не только у высших растений, но и у микроорганизмов, особенно у грибов (см. 2.2). Сидерофоры способны формировать доступные для растений Fe (ІII)-хелаты, устойчивость которых, в целом, сопоставима с этим показателем для фитосидерофоров. Однако вследствие более высокого сродства транслокатора стратегии II к фитосидерофорам последние поглощаются растениями со значительно большей скоростью, чем сидерофоры. Скорость поглощения сидерофоров и фитосидерофоров различается на два-три порядка. Снижению конкуренции между фитосидерофорами и сидерофорами за железо способствует пространственное разделение зон корня, активно участвующих в выделении и поглощении фитосидерофоров, и зон с высокой колонизацией бактерий.
Влияние внешних факторов
Механизмы стратегии II менее зависимы от величины pH среды, чем механизмы стратегии I. Этим объясняется несколько большая устойчивость злаков к хлорозу при их выращивании на карбонатных почвах. Однако в естественных условиях эффективность механизмов стратегии II может быть также невысокой, в частности, вследствие быстрого разложения фитосидерофоров микроорганизмами ризосферы, особенно при повышенных температурах. Кроме того, эффективность хелатирующего действия фитосидерофоров зависит от содержания в почве фосфора. В опытах с 59Fe установлено, что с возрастанием в карбонатной почве содержания фосфатов снижается мобилизационная в отношении железа функция фитосидерофоров.